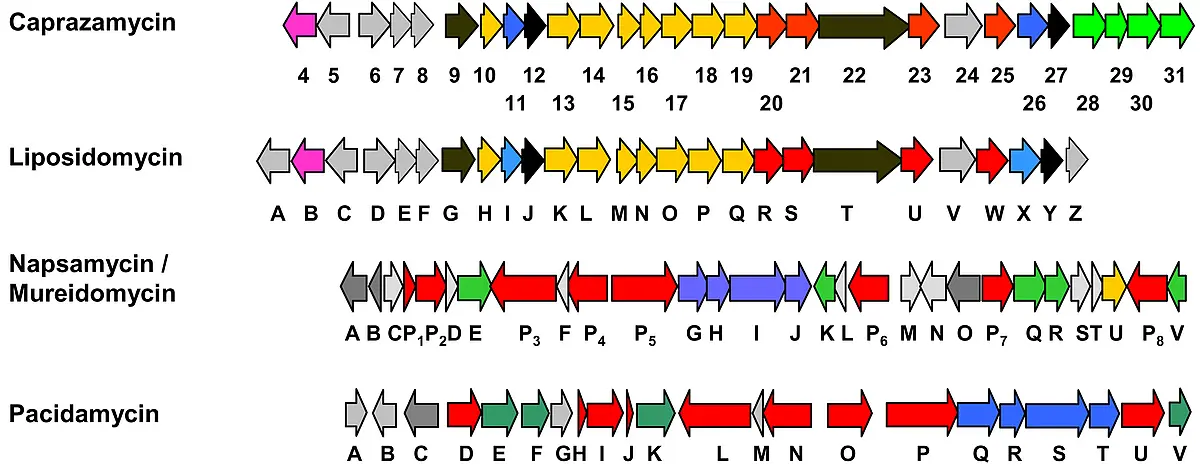
Heterologous expression of the CPZ gene cluster resulted in production of CPZ aglycones lacking the permethylated L-rhamnose moiety. Genes for the biosynthesis of the dTDP-L-rhamnose were identified elsewhere on the genome and were assembled to the CPZ gene cluster (20). The heterologous producer of the resulting gene cluster readily accumulated CPZs. A deletion mutant of the acyl-transferase cpz21 led to production of simplified liponucleoside antibiotics, the hydroxyacyl-caprazols, lacking the deoxysugar and the 3-MG moiety (17).
Analysis of the LPM gene cluster identified a sulfotransferase LpmB likely to be responsible for sulfation of the LPMs at the 2’’-position of the aminoribose. Surprisingly, an orthologue (cpz4) was identified next to the CPZ gene cluster. Investigations on the genuine CPZs producer led to identification of new, sulfated CPZ not described before (18). Cpz4 was expressed, purified and biochemically characterized (16). This enzyme is independent of 3’-phosphoadenosine 5’-phosphosulfate (PAPS) but belongs to the poorly investigated class of arylsulfate sulfotransferases (ASSTs). Kinetic investigations confirmed that the reaction proceeds by a ping pong bi-bi mechanism. The preferred substrates were CPZs, CPZs aglycones and hydroxyacyl-caprazols, demonstrating a low substrate specificity of Cpz4.
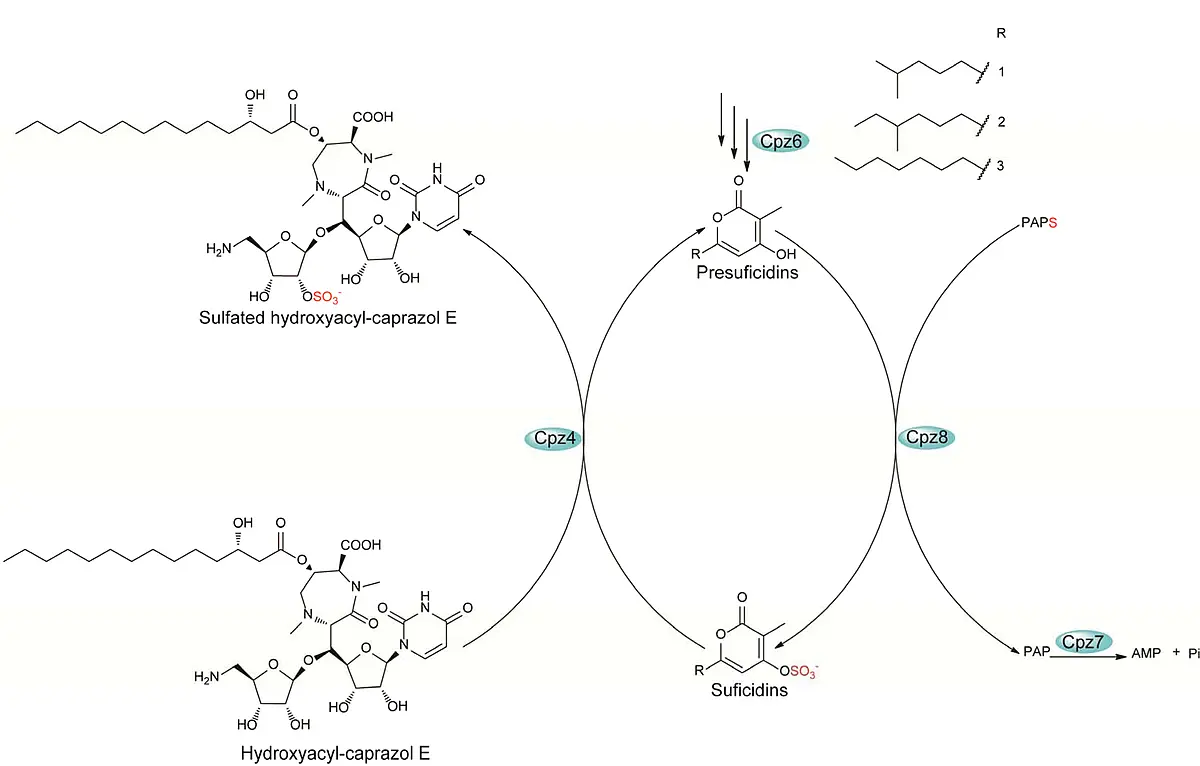
To investigate sulfation of CPZs in more detail, the proteins Cpz5-Cpz8 were characterized biochemically. A type III polyketide synthase (Cpz6) is employed in the biosynthesis of a group of new triketide pyrones that are subsequently sulfated by an unusual PAPS-dependent sulfotransferase (Cpz8) to yield phenolic sulfate esters which serve as sulfate donors for a PAPS-independent arylsulfate sulfotransferase (Cpz4) to generate sulfated caprazamycins. This finding is the first demonstration of genuine sulfate donors for an arylsulfate sulfotransferase (ASST) and the first report of a type III PKS to generate a chemical reagent in bacterial sulfate metabolism (Tang et al., 2013, Nature Chem. Biol. 9(10):610-5).