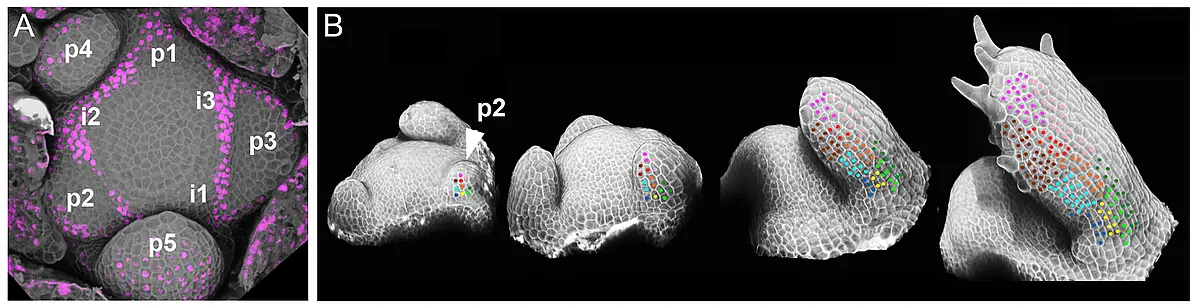
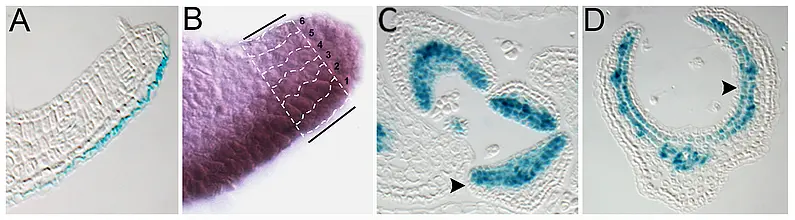
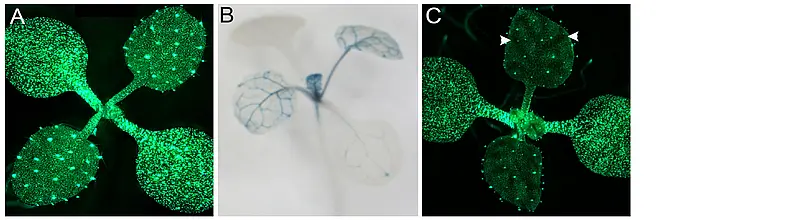
The formation of stable, precisely defined boundaries between two distinct cell fates is a fundamental feature of plant and animal development. Such cell fate boundaries coordinate the differentiation and growth of the tissue or organ. In this regard, development of flat leaf architecture poses an unusual and mechanistically challenging problem; namely, how to create a stable adaxial-abaxial (top-bottom) boundary within the plane of a long and wide, but shallow, structure? Adaxial-abaxial polarity is based on an intricate gene regulatory network with several highly conserved transcription factors that promote either adaxial or abaxial fate at its core. These are expressed in complementary domains on the top and bottom side of the developing organ, respectively. We have shown that the positional information needed to define these domains is provided in part by small RNAs that reminiscent to classical morphogens generate sharply defined domains of target gene expression through an intrinsic, threshold-based readout of their mobility gradients (Skopelitis et al., 2017).


{kind=link}