FACS
Description
Welcome to the facility for Flow Cytometry and Fluorescence Activated Cell Sorting (FACS) at the Center for Plant Molecular Biology (Zentrum für die Mokularbiologie der Pflanzen) at the University of Tübingen. Flow cytometry is a method for analysing particles in a stream of liquid and in particular, our applications are primarily based on the analysis of fluorescence. Light (photons) can be absorbed by certain molecules leading to excitation of an electron from its ground state to an excited state (an electron orbit of higher energy). In certain instances, this absorbed energy is re-emitted as light (photons) of a longer wavelength as the electron returns to the ground state; this is called fluorescence. As long as one is able to analyse their samples in solution, elicit fluorescence and maintain a particle much smaller than that of the nozzle size, one can use flow cytometric methods for applicable analyses. In addition to the measurement of fluorescence, one can get a rough estimate of particle size and particle granularity or surface differences. These attributes are independent of fluorescence. Common applications in plant science have been the measurement of DNA content for determining the phase of the cell cycle, measuring DNA content calculating the ploidy level and counting chromosomes by using fluorescent dyes that are specific to DNA. Another application has been the use of cell line specific GFP markers for the isolation of specific cell populations (e.g. quiescent root cells) by FACS followed by analysis of RNA or potentially protein content. Protein-protein interactions can also be effectively analysed by fluorescence using BiFC techniques when combined with flow. Since no microscope function associated with this particular method (non-imaging cytometry), the determination of sub-cellular compartmentalization must be made independently of the FACS treatment, either before or afterwards. Our MoFlo XDP is capable of currently detecting fluorescence excited by blue laser (488 nm), a yellow-green laser (561 nm) and either a violet laser (405 nm or 445 nm) or UV-laser (375 nm). Our CytoFLEX is equipped with a single 488 nm laser and 4 detection channels. We predominately work with plant material.
MoFlo at ZMBP

ZMBP Filter Set-up (click to view current options).
click for blue 488 path channels,
{kind=link}
click for green 561 path channels,
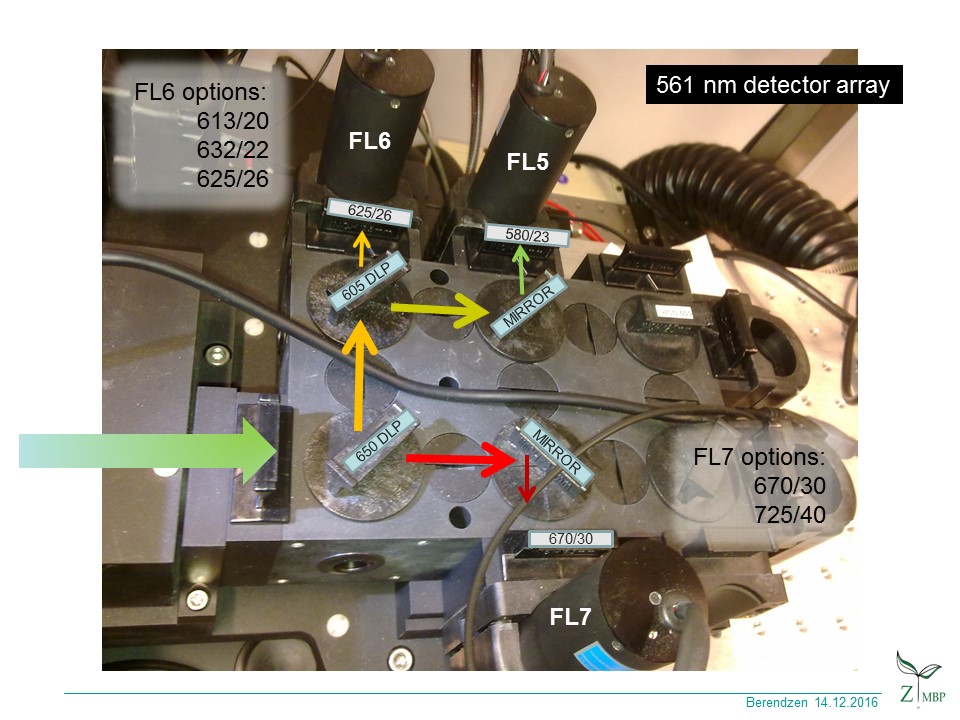{kind=link}
click for 405/375 path channels.
{kind=link}
Our FACS machine is a MoFlo (Modular Flow) XDP model from Beckman Coulter, giving us a needed flexibility required for working with plant cells. We possess 70uM, 100uM, 150uM (and 200uM) nozzles. All of these nozzles have been successfully tested with our current set-up. Our present configuration uses an OBIS 1 - 100 mW 488nm blue laser, an OBIS 1 - 100 mW 561 nm green laser for excitation, and either an OBIS UV 375nm (1 to 50 mW) or OBIS violet 405nm (1 to 100mW) or OBIS violet 445nm laser (1 to 300mW).
The fluorescence is detected through a series of dichromatic mirrors and filters which give us crude spectral data. In principle, we have filters for green, yellow, orange, red, infra-red light. Thus, with our current filter/mirror set we can detect and easily distinguish GFP/YFP and RFP/mCherry fluorescence. We are open to special applications or other filter arrangements depending on the project and demands as we have a fully modular system.
Sorting (FACS) and/or multicolor Analysis
The MoFlo XDP (BC) allows sorting of up to 4 decisions during a run, with the option to collect abort events as well. For single cell sorting events, the machine is capable of sorting into 96, 384 and 1596 (optimal alignment) well plates. Furthermore, it is possible to create user defined sorting patterns on slides, into plates, tubes, etc.. Use of the XDP requires a principle operator at all times.
Analysis (only)
Our facility has a CytoFLEX (BC) with a single 488 nm laser and 4 band passes: FL1 (525/40), FL2 (585/42), FL3 (610/20), FL4 (670/20). This is ideal for GFP/YFP, but mRFP and mCherry can also be detected. Chlorophyll fluorescence is also resolvable. After training, you are allowed to use the CytoFLEX at any time. Trouble shooting and problems should be reported to a principle Operator as soon as possible.
Samples
The XDP accepts only 5mL FACS tubes.
The CytoFLEX accommodates 5 mL FACS tubes, 2.0 and 1.5 ml Eppendorf tubes.
Biological Safety Level
Our facility is strictly S1 (BL1). No exceptions are allowed.
Note: Typically, as most mammalian cell lines carry viral particles or are generated with retroviruses, even if they are S1 (BL1), we have to consider them S2 (BL2). You can still contact us though and we will have a look; make sure you attach the appropriate data sheet(s).
Usage / Services
The MoFlo XDP and CytoFLEX is currently available to all members of the University of Tübingen, with priority given to those who are members of the ZMBP. If you wish to use the MoFlo, please contact me below. We are running an open schedule with a first-come-first-serve policy, advance bookings can be arranged for special projects (please use the email address listed below). Please inspect the User Agreement before coming. Also please read the section above about our Biological Safety Level policy.
Service Agreement and Pricing: download here
Software
We currently use Summit 5.5 software for acquisition and Summit 5.5 for the XDP. You may of course take and process your data yourself with your software of choice. The CytoFLEX comes with its own software: CytExpert. Access to FlowJo or FCS Exypress can also be provided when doing analysis "in house".
Sheath Fluid
Specific questions about sheath fluids should be personally directed to the principle operator. Typically the MoFLo XDP uses PBS [pH 7.0] and the CytoFLEX pure mili-Q purified water.
Your Data
Your data is your intellectual property. We will keep archives of the cytometry files and the set-up parameters for at least 10 years. As a user though, you are required to keep track of your samples. We will assist as much as possible if desired though. Therefore, no data or outcomes will be shared with anyone at any time without Your permission.
Flow requests, appointments, questions
facs (a) zmbp.uni-tuebingen.de
Core Manager/contact information
ZMBP - Central Facilities Kenneth W. Berendzen, Dr. rer. nat.* Auf der Morgenstelle 32 D-72076, Tübingen, Germany Office: Tel. +49-(0)7071-29-78721 Lab: Tel. +49-(0)7071-29-78715 |  |
Authorized Users
Only individuals authorized to use the MoFlo XDP are allowed to do so without constant supervision from the Principle Operator. *persons who should be addressed first. I will provide training to long term projects for in-house personnel. In this way you can run your applications themselves, but don't worry, continued support and supervision will be provided.
Current Principle Operators:
Kenneth Berendzen*
Education:
If you are interested in learning about how FACS and flow cytometry technology works and you are a Masters student, there is a course avaliable called: “Modern Methods/Introduction in Flow Cytometry”. Look for Durchflusszytometrie in Alma under Bio-ZMBP or Bio-CIB-215. In addition, you can perform lab rotations, practical courses, as well as do your Bachelor and Master thesis in the laboratory of Dr. Kenneth Berendzen. Click on the link to view his reserach interests. Topics usually include Flow Cytometry within a plant molecular-genetic framework.
Facility Publications:
2025
2024
2023
Plant J. 2023 Dec;116(6):1633-1651. doi: 10.1111/tpj.16451. Epub 2023 Sep 2.
2022
2021
A biosensor for the direct visualization of auxin. Nature. 2021 Apr;592(7856):768-772. doi: 10.1038/s41586-021-03425-2. Epub 2021 Apr 7.
Endoplasmic reticulum membrane receptors of the GET pathway are conserved throughout eukaryotes. Proc Natl Acad Sci U S A. 2021 Jan 5;118(1):e2017636118. doi: 10.1073/pnas.2017636118. Epub 2020 Dec 21.
Characterization of a Plant Nuclear Matrix Constituent Protein in Liverwort. Front Plant Sci. 2021 May 7;12:670306. doi: 10.3389/fpls.2021.670306. eCollection 2021.
2020
KINESIN-12E regulates metaphase spindle flux and helps control spindle size in Arabidopsis. The Plant Cell. 17 November 2020, koaa003,doi: /10.1093/plcell/koaa003
Isolation of Lineage Specific Nuclei Based on Distinct Endoreduplication Levels and Tissue-Specific Markers to Study Chromatin Accessibility Landscapes. Plants (Basel). 2020 Nov 3;9(11):E1478. doi: 10.3390/plants9111478.
Marchantia TCP transcription factor activity correlates with three-dimensional chromatin structure. Nat Plants. 2020 Oct;6(10):1250-1261. doi: 10.1038/s41477-020-00766-0.
(publication) Malik WA, Marco-Llorca C, Berendzen K, Piepho HP. Choice of link and variance function for generalized linear mixed models: a case study with binomial response in proteomics. Communications in Statistics-Theory and Methods 49 (17), 4313-4332
(publication) Koch M, Berendzen KW, Forchhammer AK. On the Role and Production of Polyhydroxybutyrate (PHB) in the Cyanobacterium Synechocystis sp. PCC 6803. Life (Basel). 2020 Apr 22;10(4). pii: E47. doi: 10.3390/life10040047.
(publication) Slane D, Berendzen KW, Witthöft J, Jürgens G. Transcriptomic Profiling of the Arabidopsis Embryonic Epidermis Using FANS in Combination with RNAseq. Methods Mol Biol. 2020;2122:151-164. doi: 10.1007/978-1-0716-0342-0_12.
2019
(publication) Hu B, Wang N, Bi X, Karaaslan ES, Weber AL, Zhu W, Berendzen KW, Liu C. Plant lamin-like proteins mediate chromatin tethering at the nuclear periphery. Genome Biol. 2019 Apr 30;20(1):87. doi: 10.1186/s13059-019-1694-3.
(publication) Wallmeroth N, Jeschke D, Slane D, Nägele J, Veerabagu M, Mira-Rodado V, Berendzen KW. ARR22 overexpression can suppress plant Two-Component Regulatory Systems. PLoS One. 2019 Feb 11;14(2):e0212056. doi: 10.1371/journal.pone.0212056. eCollection 2019.
(publication) Riester L, Köster-Hofmann S, Doll J, Berendzen KW, Zentgraf U. Impact of Alternatively Polyadenylated Isoforms of ETHYLENE RESPONSE FACTOR4 with Activator and Repressor Function on Senescence in Arabidopsis thaliana L. Genes (Basel). 2019 Jan 28;10(2).
2017
(acknowledgment) Triparental plants provide direct evidence for polyspermy induced polyploidy. Nakel T, Tekleyohans DG, Mao Y, Fuchert G, Vo D, Groß-Hardt R. Nat Commun. 2017 Oct 18;8(1):1033. doi: 10.1038/s41467-017-01044-y.
(publication) Zhu W, Hu B, Becker C, Doğan ES, Berendzen KW, Weigel D, Liu C. Altered chromatin compaction and histone methylation drive non-additive gene expression in an interspecific Arabidopsis hybrid. Genome Biol. 2017 Aug 22;18(1):157. doi: 10.1186/s13059-017-1281-4.
(publication) Orlando de Lange, Niklas Schandry, Markus Wunderlich, Kenneth Wayne Berendzen and Thomas Lahaye. Exploiting the sequence diversity of TALE-like repeats to vary the strength of dTALE-promoter interactions. Synthetic Biology, Volume 2, Issue 1, 1 January 2017.
2016
(publication) Durka, W., Michalski, S. G., Berendzen, K. W., Bossdorf, O., Bucharova, A., Hermann, J.-M., Hölzel, N. and Kollmann, J. (2016), Genetic differentiation within multiple common grassland plants supports seed transfer zones for ecological restoration. J Appl Ecol. doi:10.1111/1365-2664.12636
(publication) Conserved and unique features of the homeologous maize Aux/IAA proteins ROOTLESS WITH UNDETECTABLE MERISTEM 1 and RUM1-like 1. Zhang Y, Marcon C, Tai H, von Behrens I, Ludwig Y, Hey S, Berendzen KW, Hochholdinger F. J Exp Bot. 2016 Feb;67(4):1137-47. doi: 10.1093/jxb/erv519. Epub 2015 Dec 15.
2015
(publication) The Elucidation of the Interactome of 16 Arabidopsis bZIP Factors Reveals Three Independent Functional Networks. Llorca CM, Berendzen KW, Malik WA, Mahn S, Piepho HP, Zentgraf U. PLoS One. 2015 Oct 9;10(10):e0139884. doi: 10.1371/journal.pone.0139884. eCollection 2015.
(publication) Cooperative action of the paralogous maize lateral organ boundaries (LOB) domain proteins RTCS and RTCL in shoot-borne root formation. Xu C, Tai H, Saleem M, Ludwig Y, Majer C, Berendzen KW, Nagel KA, Wojciechowski T, Meeley RB, Taramino G, Hochholdinger F. New Phytol. 2015 Apr 22. doi: 10.1111/nph.13420
2014
(publication) Cell type-specific transcriptome analysis in the early Arabidopsis thaliana embryo. Slane D, Kong J, Berendzen KW, Kilian J, Henschen A, Kolb M, Schmid M, Harter K, Mayer U, De Smet I, Bayer M, Jürgens G. Development. 2014 Nov 19. pii: dev.116459.
(publication) Diversity of stability, localization, interaction and control of downstream gene activity in the Maize Aux/IAA protein family. Ludwig Y, Berendzen KW, Xu C, Piepho HP, Hochholdinger F. PLoS One. 2014 Sep 9;9(9):e107346.
2012
(publication) Screening for in planta protein-protein interactions combining bimolecular fluorescence complementation with flow cytometry. Berendzen KW, Böhmer M, Wallmeroth N, Peter S, Vesić M, Zhou Y, Tiesler FK, Schleifenbaum F, Harter K. Plant Methods. 2012 Jul 12;8(1):25.
(publication) Molecular interactions of ROOTLESS CONCERNING CROWN AND SEMINAL ROOTS, a LOB domain protein regulating shoot-borne root initiation in maize (Zea mays L.). Majer C, Xu C, Berendzen KW, Hochholdinger F. Philos Trans R Soc Lond B Biol Sci. 2012 Jun 5;367(1595):1542-51
2011
(acknowledgement) Multivesicular bodies mature from the trans-Golgi network/early endosome in Arabidopsis. Scheuring D, Viotti C, Krüger F, Künzl F, Sturm S, Bubeck J, Hillmer S, Frigerio L, Robinson DG, Pimpl P, Schumacher K. Plant Cell. 2011 Sep;23(9):3463-81. doi: 10.1105/tpc.111.086918. Epub 2011 Sep 20.
(publication) Rootless with undetectable meristem 1 encodes a monocot-specific AUX/IAA protein that controls embryonic seminal and post-embryonic lateral root initiation in maize. von Behrens I, Komatsu M, Zhang Y, Berendzen KW, Niu X, Sakai H, Taramino G, Hochholdinger F. Plant J. 2011 Apr;66(2):341-53. doi: 10.1111/j.1365-313X.2011.04495.x. Epub 2011 Mar 1.
(publication) Alanine zipper-like coiled-coil domains are necessary for homotypic dimerization of plant GAGA-factors in the nucleus and nucleolus. Wanke D, Hohenstatt ML, Dynowski M, Bloss U, Hecker A, Elgass K, Hummel S, Hahn A, Caesar K, Schleifenbaum F, Harter K, Berendzen KW. PLoS One. 2011 Feb 10;6(2):e16070.
2010
(publication) Promoter specificity and interactions between early and late Arabidopsis heat shock factors. Li M, Berendzen KW, Schöffl. Plant molecular biology. 2010 May 11.
(publication) Detection of in vivo interactions between Arabidopsis class A-HSFs, using a novel BiFC fragment, and identification of novel class B-HSF interacting proteins, Li M, Doll J, Weckermann K, Oecking C, Berendzen KW, Schöffl F., Eur J Cell Biol. 2010 Feb-Mar;89(2-3):126-32. Epub 2009 Nov 27.
2009
(acknowledgment) Regulators of PP2C phosphatase activity function as abscisic acid sensors. Ma Y, Szostkiewicz I, Korte A, Moes D, Yang Y, Christmann A, Grill E. Science.2009 May 22;324(5930):1064-8. Epub 2009 Apr 30.
Disclaimer
Although the information on this page has been made as accurate as possible, no guarantee is made concerning the content on this page nor for any of the links (and subsequent websites) they reference. The reader is encouraged to directly make contact with the webmaster or Principle Operator on this page for anything of concern. All links are of an unbiased nature and are supplied for the sole purpose of information.
Updated 26.02.2021