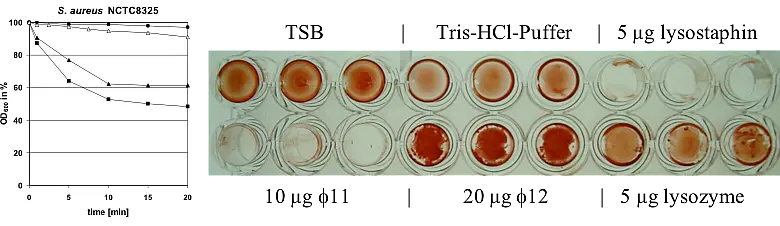
Bacterial cell wall stress response in S. aureus is mainly controlled by the two-component regulatory system VraSR that reacts to perturbation of cell wall synthesis by either cell wall-active substances, like vancomycin or daptomycin, or deregulated cell wall enzymes including MurF and Pbp2. The VraSR stimulon comprises the vraSR genes together with genes related to the cell wall metabolism of S. aureus like murZ, uppS, bacA, pbp2, sgtB and genes related to protein metabolism. We investigated the effect of the lantibiotic mersacidin, which acts by complexing the sugar phosphate head group of the peptidoglycan precursor lipid II thereby inhibiting the transglycosylation reaction of peptidoglycan biosynthesis, on inducing the cell wall stress response via VraSR. Our studies characterized the VraSR system as a stress sensing system (versus drug sensing system). We further found that mersacidin led to an extensive induction of the cell wall stress response, and in contrast to other cell wall-active antibiotics such as vancomycin, very low concentrations of mersacidin (0.15 x MIC) were sufficient for stimulon induction. The efficacy of mersacidin was not affected by an increased cell wall thickness, which is part of the VISA-type resistance mechanism and functions by trapping the vancomycin molecules in the cell wall before they reach lipid II. Therefore, the relatively higher concentration of mersacidin at the membrane might explain why mersacidin is such a strong inducer of VraSR compared to vancomycin (…more). In further studies, we characterized S. aureus VC40, a strain that shows full resistance to glycopeptides (vancomycin and teicoplanin MICs ≥32 mg/L) and daptomycin (MIC = 4 mg/L), and we could identify two amino acid exchanges in VraS amongst others by genome sequencing (…more). Transcriptomics indicated the increased expression of their respective regulons. Although not reaching the measured MIC for VC40, reconstitution of the L114S and D242G exchanges in VraS(VC40) into the susceptible parental background of S. aureus NCTC 8325 resulted in increased resistance to glycopeptides and daptomycin. The expression of VraS(VC40) led to increased transcription of the cell wall stress stimulon, a thickened cell wall, a decreased growth rate, reduced autolytic activity and increased resistance to lysostaphin-induced lysis in the generated mutant. Hence, a double mutation of a single gene locus, vraS, is sufficient to convert the vancomycin-susceptible strain S. aureus NCTC 8325 into a vancomycin-intermediate S. aureus (…more).
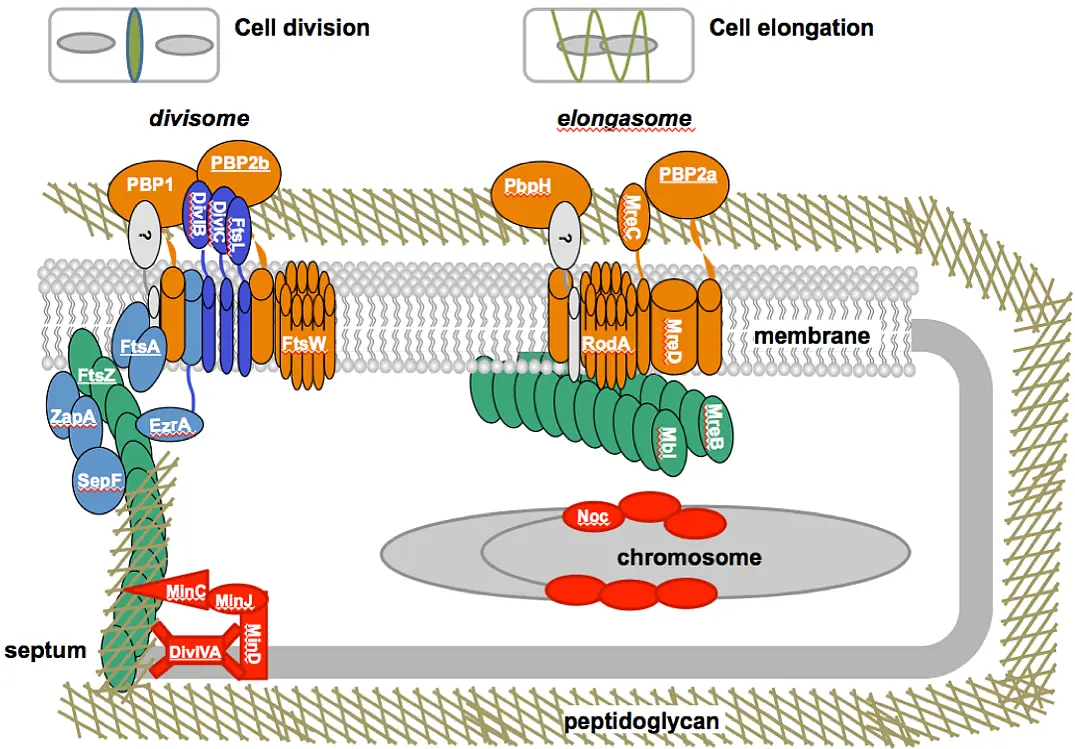
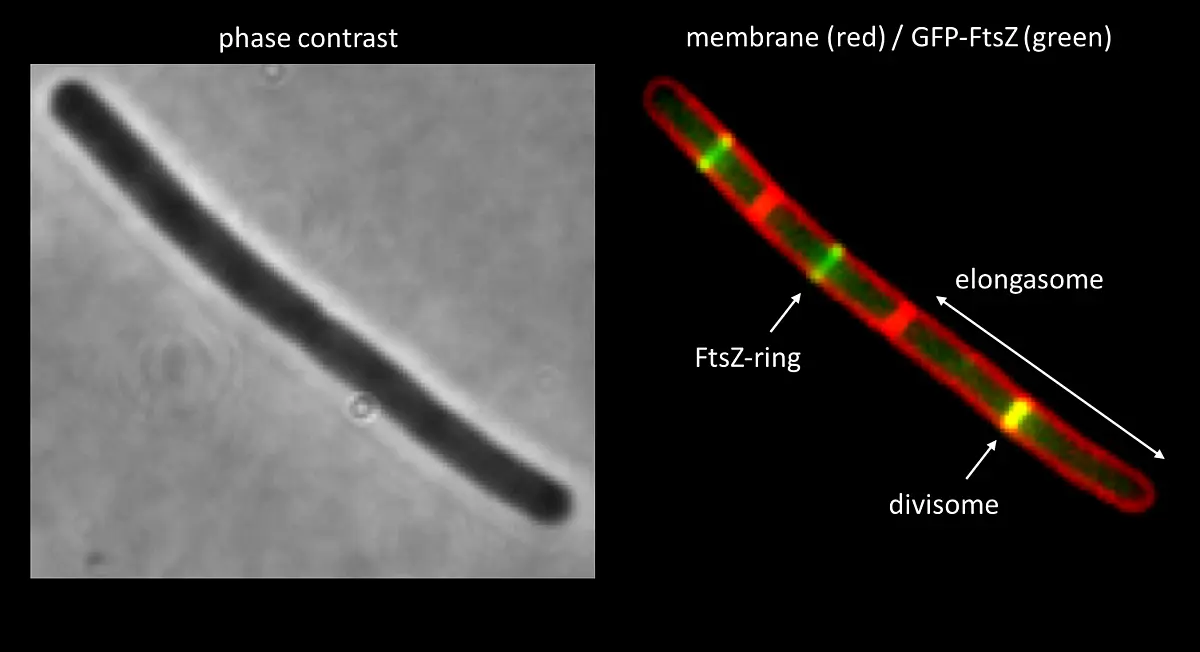
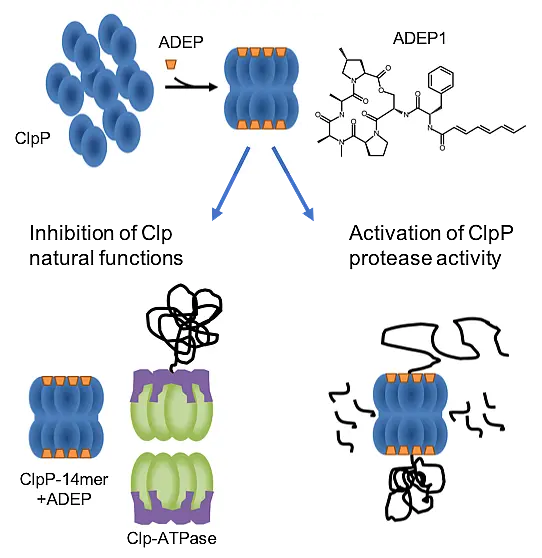

 were incubated in the presence of PknB for 60 min. A, autoradiogram; C, Coomassie stain of SDS PAGE.")