Control and activity of the Streptomyces Spore wall Synthesizing Complex SSSC
Coworkers
Nadja Steblau (PhD student), Bernd Vollmer (PhD student), Annette Latus (technician)
A major determinant of bacterial cell shape is the peptidoglycan (PG), which consists of glycan strands, cross-linked by short peptides. Bacteria control their shape by the specific localization of their cell wall synthesizing machineries. A key component of rod-shaped bacteria is the actin-like MreB, which polymerizes into filaments. MreB was shown to interact with other morphogenic proteins in rod-shaped bacteria and forms a lateral cell wall synthesis complex with MreC, MreD, PBP2, and RodA, directing incorporation of newly synthesized PG along the cylindrical part of the cell in a helical manner (White & Gober Trends Microbiol.2012. 20:74-9.
In contrast to rod-shaped bacteria which depend on MreB proteins to control their cell wall assembly, many Gram-positives of the phylum Actinobacteria grow in a different way by building their cell walls at the poles. Corynebacteria and mycobacteria do not contain mreB genes. They acquire rod shape by polarised growth, depending on the coiled-coil protein DivIVA. Similarly, streptomycetes, which do not divide by binary fission and grow by apical tip extension to form a multiply branching mycelium, also depend on DivIVA to direct this mode of apical growth (http://www.ncbi.nlm.nih.gov/pubmed/21036658). Against this background, it was a surprise that an mreB cluster comprising mreB, mreC, mreD, pbp2 and sfr (rodA) is present in S. coelicolor (http://www.ncbi.nlm.nih.gov/pubmed/10954092). Later it was shown that mreB and its paralogue mbl were dispensable for apical growth of vegetative hyphae. Instead, spore morphology was affected and the spores had lost their resistance to heat, detergent, high osmolarity, lysozyme, and vancomycin (http://www.ncbi.nlm.nih.gov/pubmed/16677297, indicating a defective spore wall. A very similar phenotype was observed for mutants defective in one of the other mre-genes (https://www.ncbi.nlm.nih.gov/pubmed/21244527). MreB and its paralogue Mbl were localized by fluorescence microscopy to the sporulation septa and the spore envelope www.ncbi.nlm.nih.gov/pubmed/21257777 .
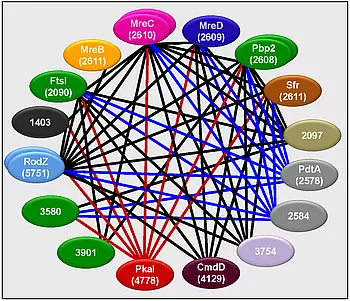
Analysis of protein-protein interaction by a bacterial two-hybrid system indicated that the S. coelicolor Mre proteins form a spore wall synthesizing complex which closely resembles the lateral wall synthesizing complex of rod-shaped bacteria (https://www.ncbi.nlm.nih.gov/pubmed/21244527). Screening of a genomic library identified several novel putative components of this complex, e.g.:
- SCO2097, an actinomycetes specific membrane protein encoded by the dcw cluster,
- SCO4129 (cmdD), previously shown to be involved in morphological differentiation
- the eukaryotic type serine/threonine kinase SCO4778 (pkaI), suggesting control of spore wall synthesis by phosphorylation
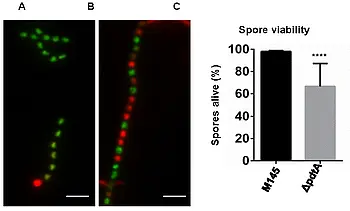
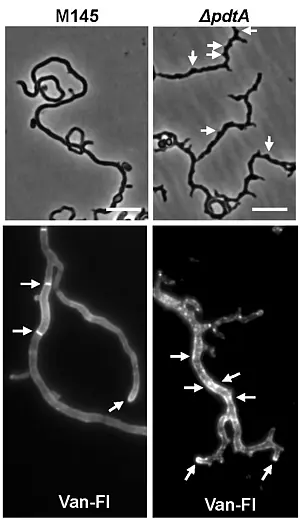
- putative cell wall glycopolymer (CWG) biosynthesis proteins (SCO2578 (pdtA) and SCO2584), indicating a close interconnection between the synthesis of PG and spore wall glycopolymers