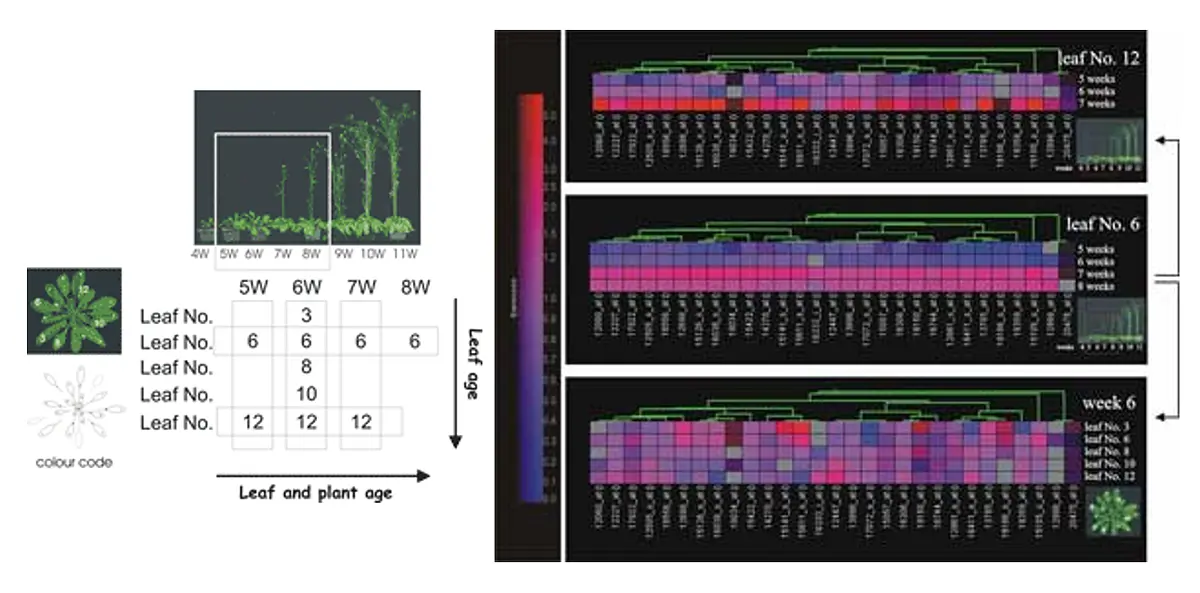
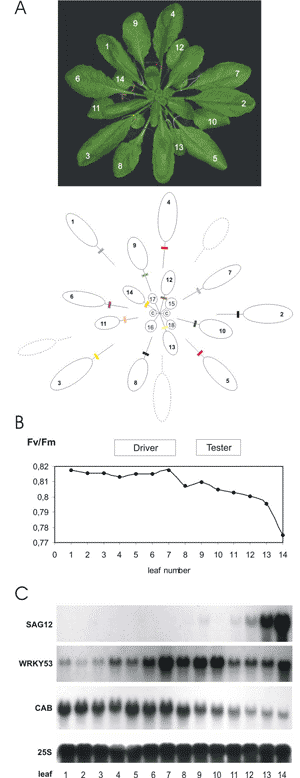
The differential expression of genes according to the age of the leaves within one rosette was also analyzed using suppression subtractive hybridization (SSH). A transcription factor of the WRKY family, WRKY53, was isolated in this screen (Hinderhofer et al., 2001).
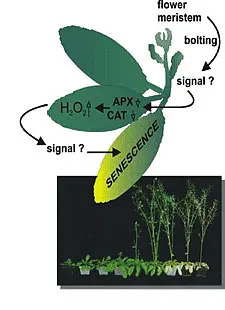
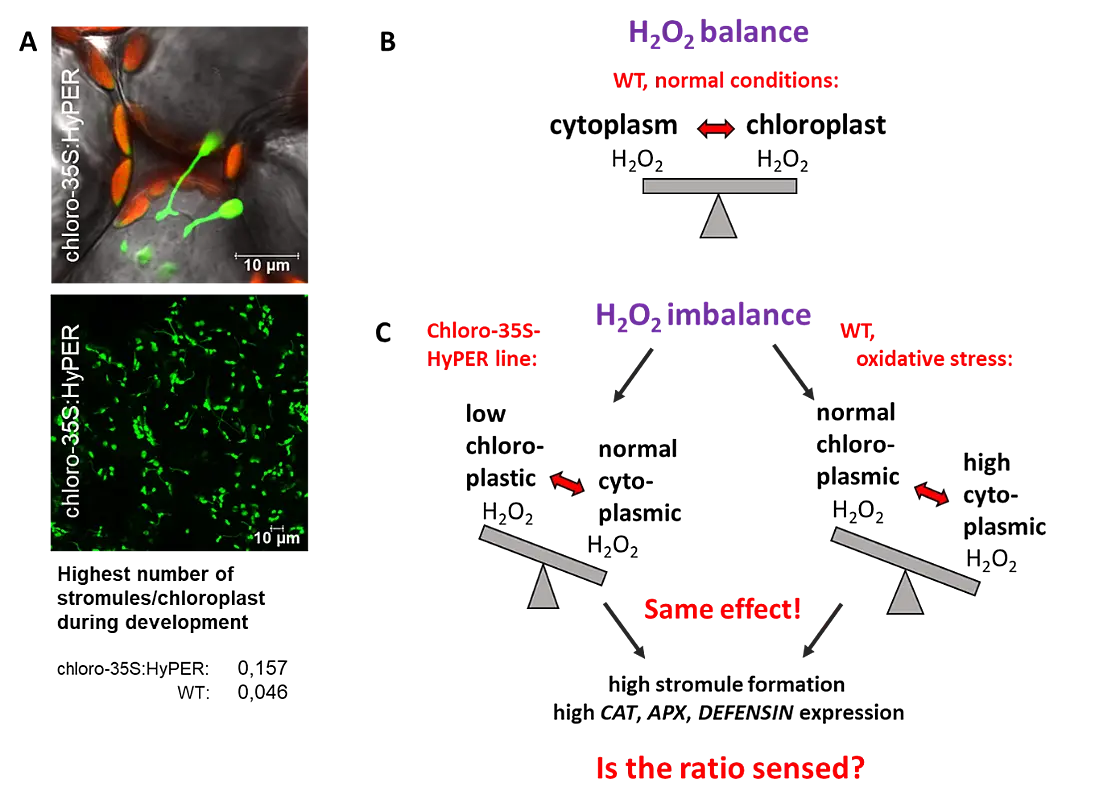
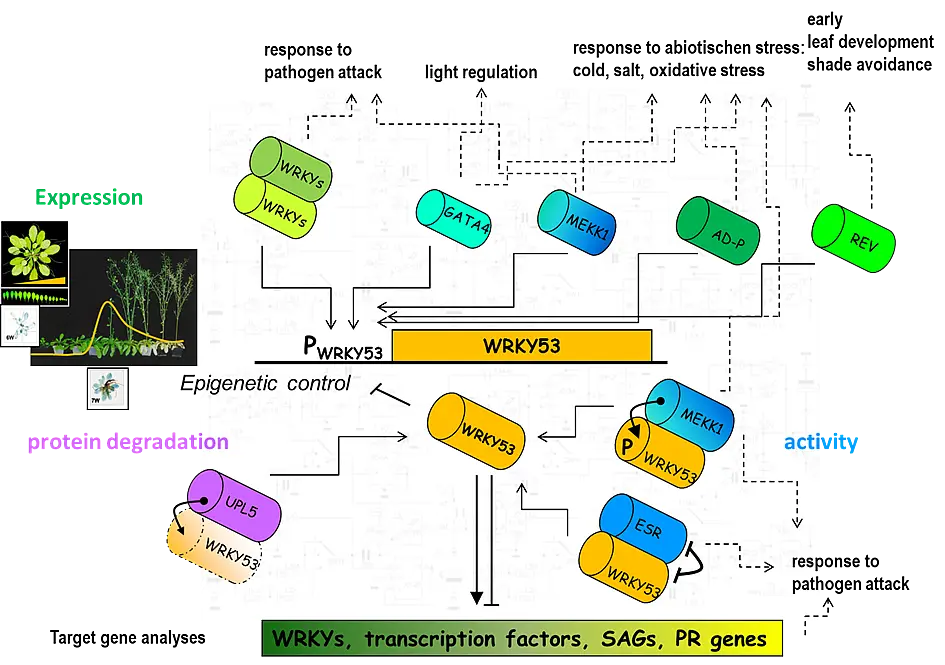
To understand the regulatory role of the senescence related WRKY53 factor, we identified targets of this transcription factor by a pull down assay using genomic DNA and recombinant WRKY53 protein. We isolated a number of candidate target genes including other transcription factors, also of the WRKY family, stress- and defence related genes, and senescence-associated genes (SAGs). WRKY53 protein could bind to these different promoters in vitro and in vivo and it could act either as transcriptional activator or transcriptional repressor depending on the sequences surrounding the W-boxes. WRKY53 can be induced by H2O2 and can regulate its own expression in a negative feed-back loop (Miao et al., 2004). Our results suggest that WRKY53 acts in a complex transcription factor signaling network regulating senescence specific gene expression and hydrogen peroxide might be involved in signal transduction.
WRKY53 is tightly regulated on different levels: i) activity, positively by phosphorylation (MEKK1) and negatively by blocking the DNA-binding (ESR/ESP), ii) gene expression which is regulated by at least four proteins that bind to the promoter of WRKY53 and iii) degradation of the protein by a HECT E3 ubiquitin ligase (Miao and Zentgraf, 2007, Miao et al., 2007, Miao et al., 2008, Miao and Zentgraf, 2010, for review see Zentgraf et al., 2010, Potschin et al., 2014).