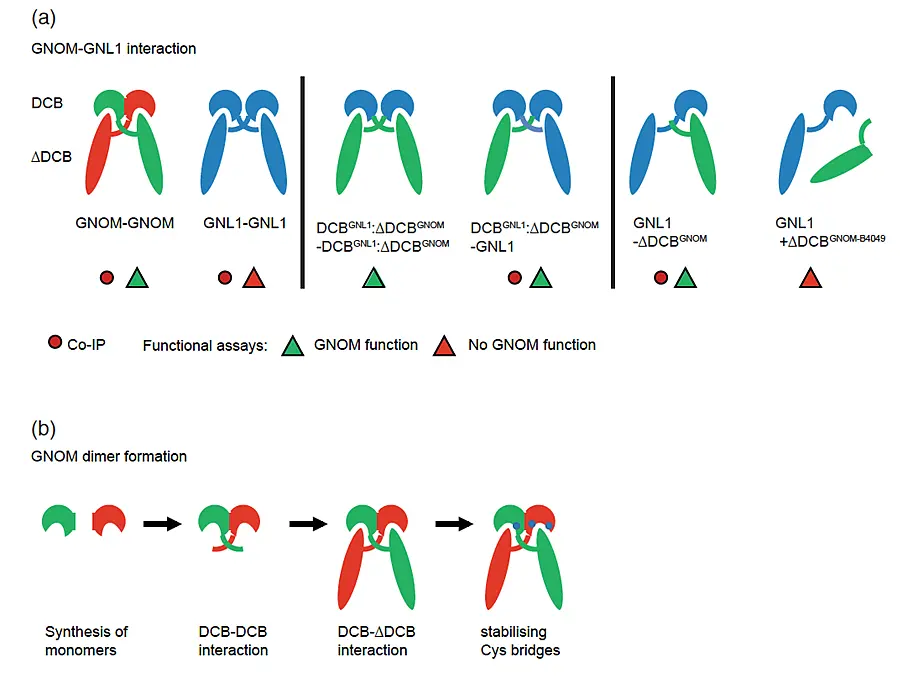
The Arabidopsis thaliana genome encodes eight 150-220kDa large ARF-GEFs that are members of a conserved eukaryotic protein family comprising two classes (GBF1, BIG1) named after their mammalian ARF-GEFs. Three proteins (GNOM, GNL1 and GNL2) are related to GBF1 acting atcis-Golgi whereas fiveproteins (BIG1-5) are related to BIG1 acting at trans-Golgi (Anders & Jürgens, 2008).
GNL1 regulates the retrograde COPI-mediated traffic from the Golgi stacks to the ER, which is the ancient eukaryotic function of the GBF1 class (Fig. 1; Richter et al., 2007). GNOM can replace GNL1 in retrograde Golgi-ER traffic. However, GNOM primarily regulates polar recycling of the auxin-efflux transporter PIN1 from endosomes to the basal plasma membrane, which is required for cell polarity and axis formation in embryogenesis, and for polar auxin transport to the root pole (Fig. 1; Geldner et al., 2003). The third paralog GNL2 plays a specific role in pollen germination and pollen-tube growth (Richter et al., 2012). The ARF-GEFs BIG1 to BIG4 jointly perform an essential function in the late-secretory pathway from the trans-Golgi network (TGN) to the plasma membrane and during cytokinesis (Fig. 1; Richter et al., 2014). Although acting at different (endo)membranes, all Arabidopsis ARF-GEFs appear to activate GTPases of the same ARF1 class which are 99% identical (Singh et al., 2018).
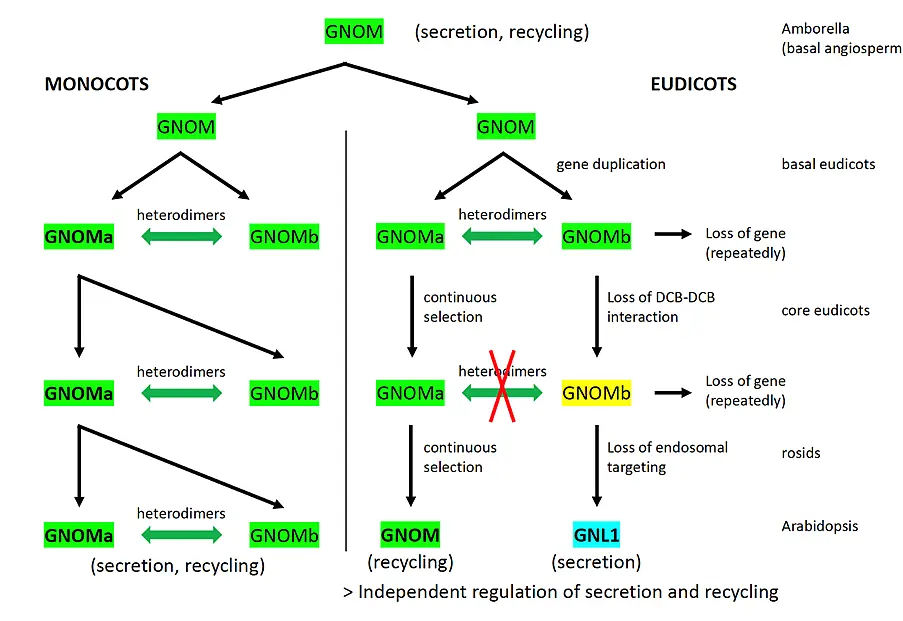
In evolution, GNOM was the “ancient plant” GBF1-type ARF-GEF and was bi-functional, mediating polar recycling and Golgi-ER retrograde traffic (Fig. 3; Singh, Lauster and Huhn et al., 2025). GNL1 only arose by duplication and diversification during eudicot evolution, although GNL1 has the GBF1-equivalent role in trafficking.