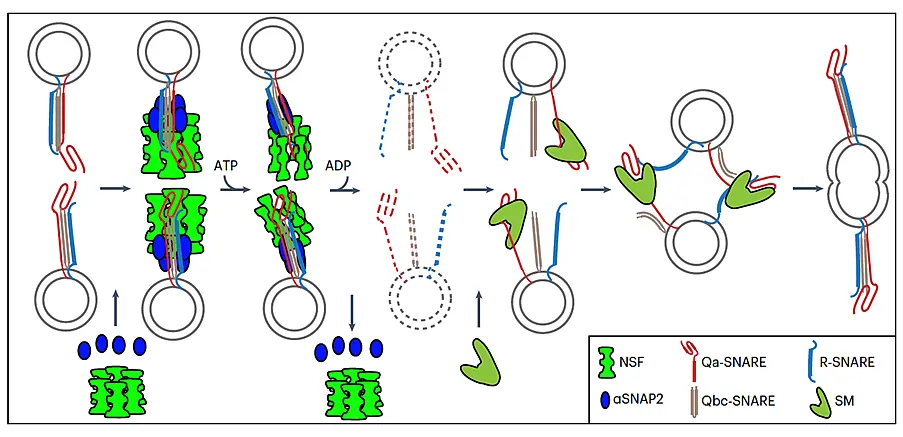
Figure 6. Trafficking of cis-SNARE complexes during cytokinesis (model).
Two different types of cytokinetic cis-SNARE complexes are assembled on the ER, recruited into COPII vesicles and passed on to the Golgi stack/TGN. At the TGN, the cis-SNARE complexes are incorporated into AP1/CCV vesicles for delivery to the division plane. Following their disassembly by NSF ATPase, monomeric Qa-SNARE KNOLLE is assisted by SM protein KEULE in the formation of trans-SNARE complexes mediating fusion of adjacent vesicles during cell-plate formation and expansion (Karnahl & Park et al., 2017).
Cell-plate formation involves newly-synthesised proteins including KNOLLE as well as proteins endocytosed from the plasma membrane which are recycled back to the plasma membrane in interphase. During cytokinesis, however, recycling of endocytosed PM proteins does not occur. These proteins are instead re-directed along a late-secretory pathway dependent on ARF-GEFs BIG1-4 to the plane of cell division (Richter et al., 2014).
Before vesicle fusion during cell-plate formation, the cis-SNARE complexes are disassembled by the action of AAA ATPase NSF and its co-factor aSNAP2 (Fig. 7; Park et al., 2023). Monomeric KNOLLE is then prevented from adopting an inactive "closed" state by interaction with the regulatory Sec1p/Munc18 (SM) protein KEULE (KEU), which promotes trans-SNARE complex formation between neighbouring membrane vesicles (Park et al., 2012).