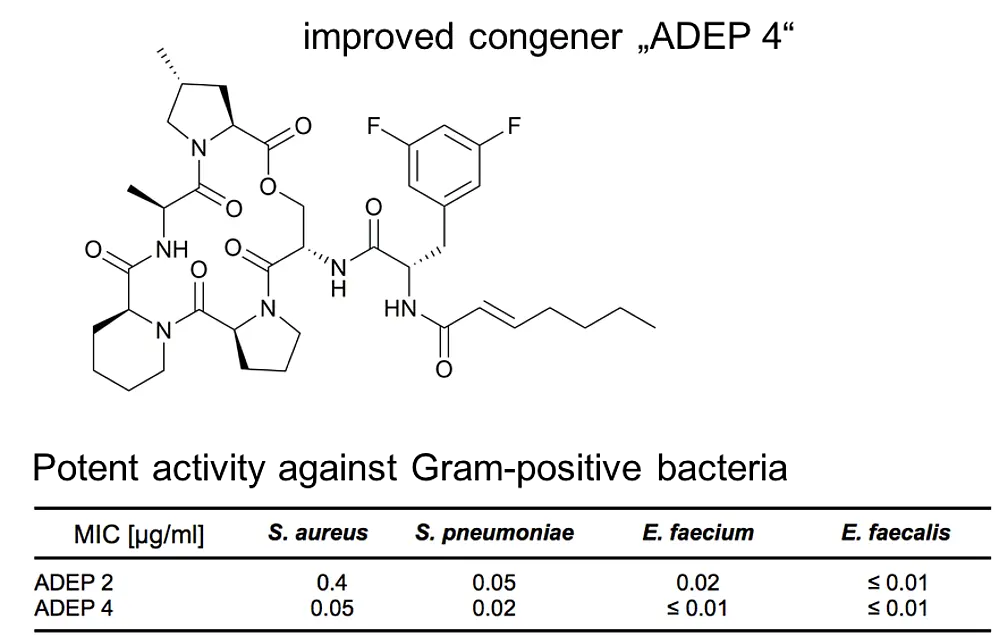
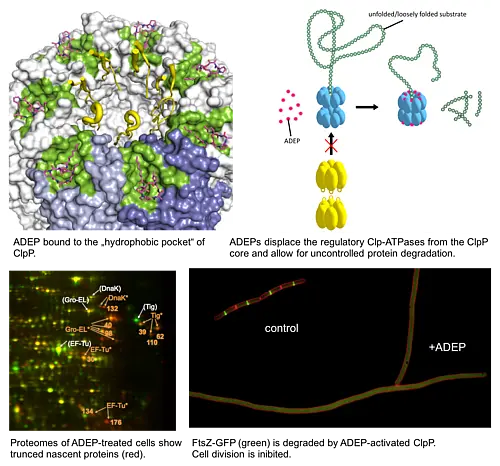
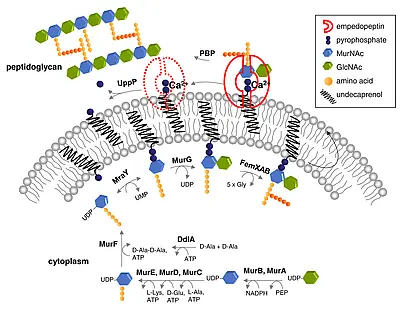
ADEP also stands apart from other antibiotics in terms of mode of action; it targets and dysregulates the proteolytic component of the bacterial Clp protease system, ClpP (...more).The bacterial Clp protease system plays an important role in maintaining protein homoeostasis within bacterial cells and in directing cellular differentiation and development programs. It is essential for bacterial virulence and serves as drug target of several recently discovered antibacterial natural products. ClpP is the conserved proteolytic core of the bacterial Clp protease complexes. For protein degradation, ClpP strictly depends on cognate Clp-ATPase partners. Without Clp-ATPases ClpP is only capable of digesting small peptides. Clp-ATPases act as a safety measure to ensure that ClpP degrades only such proteins that are destined for degradation (...more).
ADEP antibiotics unleash ClpP from these regulatory constrains. Upon binding of ADEP, ClpP sets out to degrade essential bacterial proteins, among those unstructured proteins and nascent polypeptide chains as they emerge from the ribosome. ADEP-treated cells die by self-digestion (...more). In firmicutes, such as staphylococci, streptococci, enterococci and bacilli the cell division regulator FtsZ proved to be especially sensitive to degradation by ADEP-activated ClpP and in these bacteria cell division inhibition is the primary cause of ADEP-mediated death (...more). FtsZ is also degraded in Wolbachia endobacteria, endosymbionts which pathogenic filarial worms require for reproduction (...more). In contrast, ADEP kills mycobacteria by inhibiting the indispensable natural functions of the Clp protease system (...more).
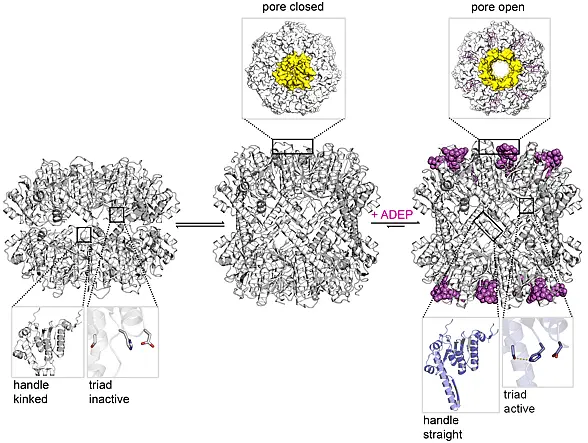
ADEP deregulates ClpP by extensive conformational control. ClpP is a 14-mer serine protease, with 14 catalytic centres shielded within its central proteolytic cavity. Small entrance pores prevent access of proteins to the proteolytic compartment. Upon binding of ADEP conformational rearrangements are set in motion. One consequence is that the gated pores widen, now allowing entry of proteins that the cell needs for survival. A second consequence is that ClpP is stabilised in the state where the catalytic triade adopts the conformation required for catalysis (...more). ADEP is the first antibiotic described to deregulate and over-activate its target; it turns a well-controlled bacterial protease into a deadly protein shredder.