Biology of Streptomyces plasmids: Mechanism of conjugative DNA-transfer in antibiotic producing streptomycetes
Biology of Streptomyces plasmids: Mechanism of conjugative DNA-transfer in antibiotic producing streptomycetes
As producers of antibiotics, actinomycetes are regarded as the natural source and reservoir of antibiotic resistance genes which have been developed as part of the biosynthetic gene clusters to protect the producer from its own antibiotic. By horizontal gene transfer, the resistance determinants probably found their way into pathogenic bacteria causing major health problems.
Streptomycetes contain a plethora of mobile genetic elements including small and larger circular plasmids, huge linear plasmids of several hundred kbp in size, and Actinomycete Integrative and Conjugative Elements (AICE). These elements direct the shaping of Streptomyces genomes and might have a particular role in the evolution and diversification of biosynthetic gene clusters.
With few exceptions, Streptomyces plasmids do not encode any beneficial traits for the host cell. Even small Streptomyces plasmid, like pSVH1, (http://www.ncbi.nlm.nih.gov/pubmed/16439019) having a size of 8-15 kb and replicating via the rolling-circle mechanism are self-transmissible and encode all functions required for plasmid transfer and the mobilization of chromosomal markers (Fig.1).
Streptomyces conjugation has been studied for 60 years but its underlying molecular mechanism is widely unknown. Conjugative DNA-transfer in mycelial actinomycetes is a unique process, differing considerably from conjugation via a type IV secretion system (T4SS), which has been well studied in many gram-negative and gram-positive bacteria (http://www.ncbi.nlm.nih.gov/pubmed/19946141).
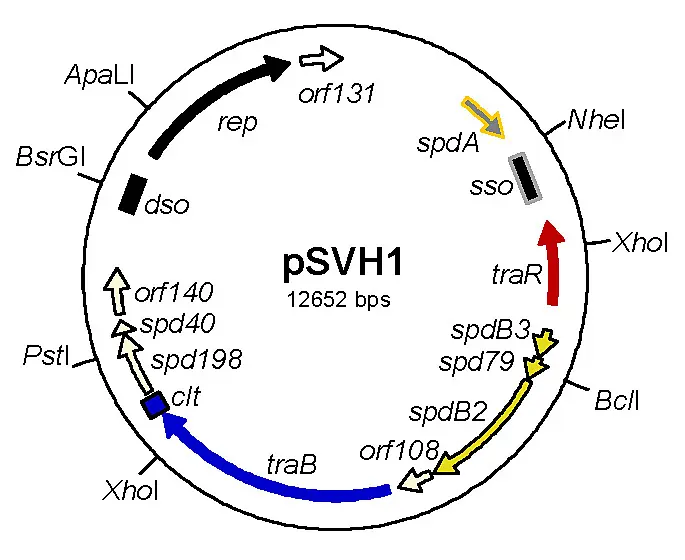
Fig. 1. Map of the conjugative plasmid pSVH1 from the chloramphenicol producer Streptomyces venezuelae. Functions involved in replication (black), regulation (red), transfer (blue) and intramycelial plasmid spreading (yellow) are indicated.
Conjugative plasmid transfer, a two-step process
In contrast to conjugation via a T4SS, Streptomyces conjugation involves the transfer of a double-stranded plasmid molecule (http://www.ncbi.nlm.nih.gov/pubmed/11679075). Under certain conditions, the Streptomyces DNA-transfer process is associated with the formation of inhibition zones, which have been named pocks. Pock structures are formed, when spores of a plasmid-carrying donor are plated with an excess of plasmid-free recipient spores. Under these conditions pocks of up to 3 mm develop in the growing mycelium (Fig.2).

{kind=link}
Fig. 2. Pock structures formed during conjugative transfer of plasmid pSVH1. S. lividans TK23 carrying the pSVH1 derivative pEB211 was streaked on a lawn of plasmid free S. lividans TK64.
These “pocks” represent inhibition zones where growth and morphological differentiation is temporary retarded and indicate areas in the mycelial lawn, where a recipient has obtained a plasmid by conjugation (https://www.ncbi.nlm.nih.gov/pubmed/27687731). Although the molecular mechanism underlying the formation of pock structures has not been elucidated, they have been interpreted as the consequence of intramycelial plasmid spreading within the recipient. This hypothesis was confirmed by the fluorescence microscopic imaging of Streptomyces conjugation. Using a eGFP-encoding derivative of the conjugative pIJ101 plasmid and a recipient strain encoding mCherry, donors, recipients and transconjugant mycelia could be differentiated (Fig.3). Observation of mating hyphae demonstrated that mating involves the lateral walls and not the primary tips, as it was previously suggested (http://www.ncbi.nlm.nih.gov/pubmed/16776656). Since mCherry fluorescence was never detected in the donor compartments making contact to the recipient, it can be concluded that conjugative transfer does not involve intermixing of the cytoplasms, but more likely proceeds via a DNA-specific translocation pore (https://www.ncbi.nlm.nih.gov/pubmed/26286483). Following the initial transfer, the newly transferred plasmid then spreads from the contact sites of donor and recipient to the older parts of the recipient mycelium across septal cross walls.
Conjugative plasmids are well adapted to the Streptomyces life style and colonise the recipient mycelium by the combination of several distinct processes: i, the plasmid propagates in the extending tip compartments and enters new hyphae at branching points, before cross walls are formed. ii, the initial conjugative transfer event is then multiplied by subsequent rounds of conjugation, since the transconjugant hyphae have been transformed into donors and can transfer the plasmid, whenever a recipient is met. iii, the development of the Spd apparatus allows the plasmid to travel across preformed cross walls into older mycelial compartments. The ability of the plasmid to spread to older parts of the recipient colony seems to be the most significant process, as demonstrated by the dramatic reduction in pock size of a spd mutant.
Moreover, plasmid spreading into “old” parts of the mycelium might be important for the dissemination of the plasmid within the environment. The older colony parts usually suffer from nutrient depletion and, as a consequence, start to differentiate into aerial hyphae, which eventually are converted into spore chains. Therefore, plasmid spreading ensures that plasmids arrive in differentiating hyphae in time and are packaged in spores.
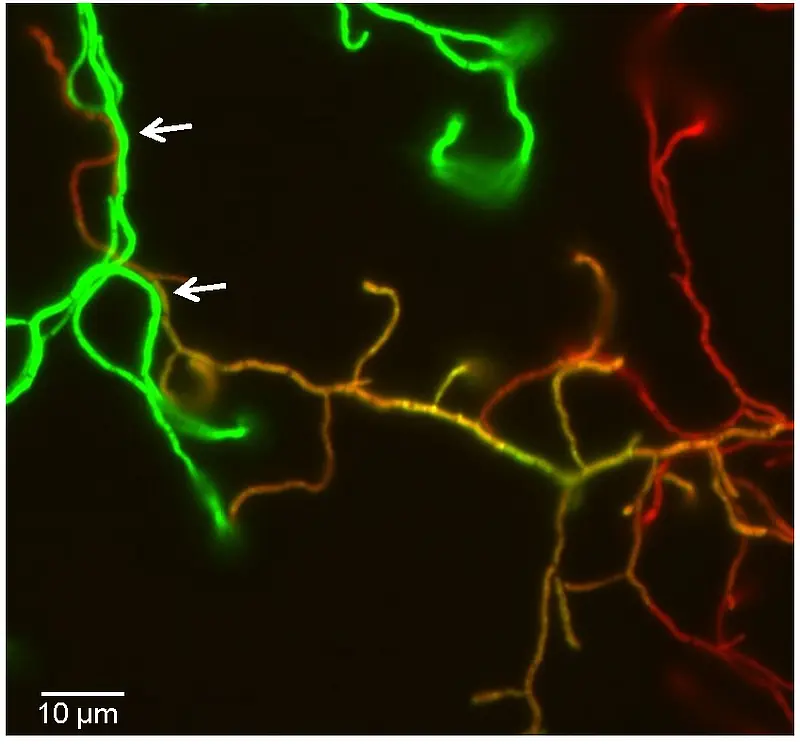
Fig. 3. Vizualization of conjugative transfer and plasmid spreading within recipient hyphae. Spores of the donor S. lividans TK23 (pLT303) (green) and the recipient S. lividans T7–mCherry (red) were mixed and plated. After 20 h of growth at 29°C cells were imaged by fluorescence microscopy. Transconjugant hyphae appear yellow in the overlay. Scale bar, 5 µm.
Initial plasmid transfer by the AAA+ ATPase TraB
A single plasmid encoded protein (TraB) is sufficient to direct the initial conjugative DNA-transfer (http://www.ncbi.nlm.nih.gov/pubmed/23082971) to the recipient. TraB proteins encoded by different Streptomyces plasmids are highly variable but resemble the septal chromosome translocator proteins FtsK/SpoIIIE in domain architecture, structure, and mode of DNA interaction. Purified TraB of plasmid pSVH1 is a DNA-dependent ATPase that is highly stimulated by dsDNA substrates (https://www.ncbi.nlm.nih.gov/pubmed/30723158). TraB monomers assemble to hexameric ring-shaped structures (and higher-order assemblies) that are able to form pores in planar lipid bilayers (Fig.4).
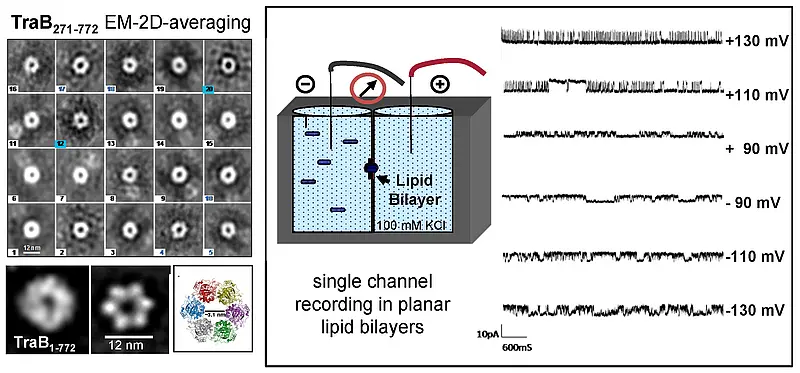
Fig.4. Structure and pore forming ability of TraBpSVH1. Electron microscopy of TraB shows a ring-shaped morphology. 2D averaging of electron microscopic images and homology modelling (template DNA-translocase domain of P. aeruginosa FtsK) of soluble TraB lacking the N-terminal membrane domain show a hexameric structure with a ~ 12 nm central channel. Single channel recordings in planar lipid bilayers revealed a pore forming ability of TraB.
TraB proteins bind to the plasmid localized cis acting locus of transfer (clt) by recognizing series of direct 8-bp repeats (TraB Recognition Sequence, TRS). Construction of various chimeric TraB proteins demonstrated that helix α3 of the c-terminal winged-helix-turn-helix (wHTH) fold determined sequence specific DNA recognition (http://www.ncbi.nlm.nih.gov/pubmed/21505418).
Model of Conjugative DNA-transfer in Streptomyces
The characterization of the pSVH1-encoded Tra and Spd proteins suggested the following model of the Streptomyces conjugative DNA-translocation system (http://www.ncbi.nlm.nih.gov/pubmed/25592263). Streptomyces plasmids do not seem to encode a system for establishing mating pairs. Therefore, conjugative DNA-transfer occurs only on solid agar during growth as substrate mycelium. Formation of multiple branching hyphae by apical tip extension provides good probability that mating partners will meet. Since Streptomyces hyphae grow by apical tip extension, the machinery for peptidoglycan remodeling is present at the hyphal tip and at new branching points. Presence of TraB probably directs fusion of the walls of the mating partners, maybe by interaction with a chromosomally encoded lytic transglycosylase.
After fusion of the PG-layer, Membrane insertion of TraB hexamers that assembled at clt sequences provides a pore structure for the DNA translocation.
TraB was shown to bind to the clt locus non-covalently without processing the plasmid (http://www.ncbi.nlm.nih.gov/pubmed/16776656). This raises the question, how TraB is able to translocate a circular DNA molecule. While during cell division the chromosome is already present at the closing septum, allowing FtsK to assemble at KOPS sequences at each chromosomal arm, TraB has to translocate the DNA across an intact membrane. Since it is highly unlikely that a circular molecule can be transported across the membrane, involvement of further DNA processing enzymes, e.g. a topoisomerase has to be postulated. Another possibility would be the fusion of the two TraB hexamers to a single channel structure releasing the circular plasmid end into the recipient.
In the recipient a vegetative cross wall traversing multi-protein complex consisting of Spd proteins is formed. By interaction with SpdB2 and other Spd proteins (http://www.ncbi.nlm.nih.gov/pubmed/17768240, www.ncbi.nlm.nih.gov/pubmed/27687731) the DNA translocase TraB is redirected to the septal crosswall and promotes plasmid transfer to the neighboring mycelial compartments. Thus, through the combined action of initial plasmid transfer and subsequent plasmid spreading large parts of the recipient mycelium are colonized by the plasmid. Since TraB not only interacts with the clt locus but also recognizes chromosomal clc sites, plasmid transfer also involves the transfer of chromosomal DNA. In mating experiments with specifically marked donor and recipient strains, we observed the high-efficient transfer of megabase-sized chromosomal DNA-fragments (or a complete donor chromosome), containing multiple Biosynthetic Gene Clusters (BGC). Apparently, the Streptomyces chromosome can exploit the plasmid-encoded transfer apparatus, thereby acting itself as a mobile genetic element.
Vector plasmids
Actinomycetes genes are often difficult to express in heterologous hosts and purification of the respective proteins in active form can be challenging. Thus, a series of stable replicating bifunctional expression vectors has been constructed to facilitate gene expression in Streptomyces strains (https://www.ncbi.nlm.nih.gov/pubmed/30191290).
Most vectors are derived from the multi-copy, temperature-sensitive Streptomyces ghanaensis plasmid pSG5, which replicates via a rolling-circle replication mechanism (RCR). To support replication in E. coli the pMB1 replicon was included. Resistance genes against thiostrepton (tsr) and kanamycin (aphII) or apramycin (aac(3)IV) allow selection in Streptomyces and E. coli. Stable replication in Streptomyces was achieved by placing all genes in the same orientation as rep and by inserting the single-stranded origin sso for lagging strand replication. Inducible gene expression is mediated by either the PtipA promoter or the PT7 promoter (for use with S. lividans T7, constructed by J. Altenbuchner). The PSF14 promoter is constitutively active in E. coli and Streptomyces. Thus, Streptomyces and E. coli colonies carrying pGM1192 show a pink colour, even without irradiation with blue light (https://www.ncbi.nlm.nih.gov/pubmed/30191290). Some vectors, like pGM1202 are equipped with the oriT fragment of plasmid RP4 and can be mobilized to streptomycetes by intergeneric conjugation. His-tag and Strep-tag II encoding sequences allow purification of the proteins via affinity chromatography. Using these plasmids several Streptomyces proteins that could not be produced in E. coli were purified from S. lividans. Some of the plasmids are available from Addgene (https://www.addgene.org/).
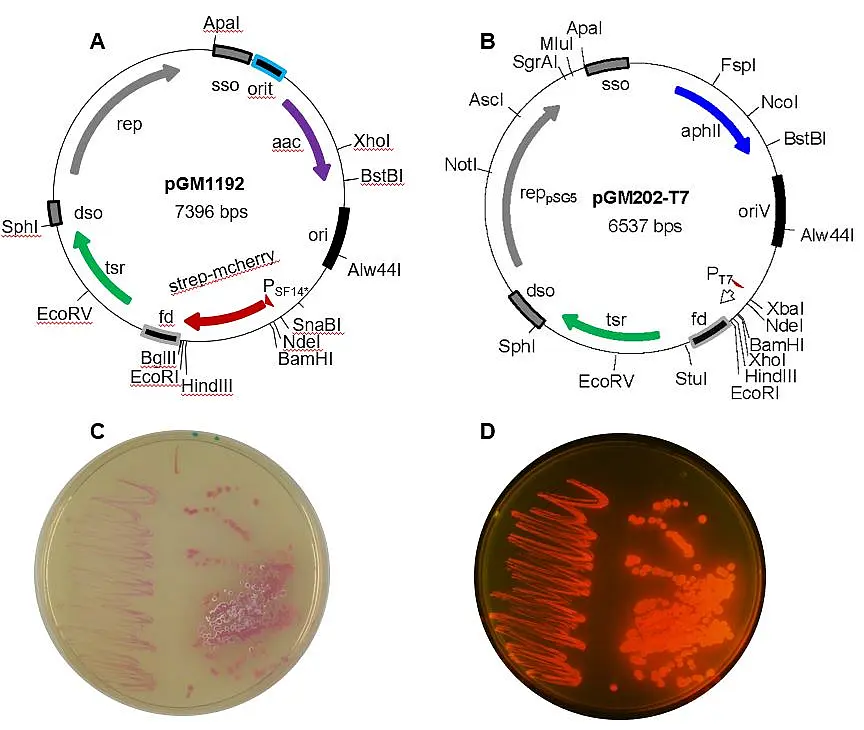
Fig. 5. Restriction maps of two bifunctional Streptomyces vectors based on the minimal replicon of pSG5. pGM1192 (A) carries a codon optimized mCherry gene under control of the strong PSF14* promoter of the actinophage I19, while pGM202T7 (B) contains the PT7 promoter. Colonies of E. coli (left side) or S. lividans (right side) carrying pGM1192 appear pink/red on LBApra plates, without (C) and with UV irradiation (D).
Table 1. Characteristics of some bifunctional Streptomyces (expression) vectors
vector | characteristics |
pGM190 | tsr, aphII, to terminator PtipA, RBS, fd terminator, sso, repts |
pGM190oriT | tsr, aphII, oriT, to terminator PtipA, RBS, fd terminator, sso, repts |
pGM191.1N | tsr, aphII, to terminator PtipA, RBS, N-terminal Strep-tagII-eGFP, fd terminator, sso, repts |
pGM202 | tsr, aphII, to terminator PtipA, RBS, C-terminal His6-tag, fd terminator, sso, repts |
pGM202-T7 | tsr, aphII, PT7, RBS,N- and C-terminal His-tags, fd terminator, sso, repts |
pGM202-T7-mcherry | tsr, aphII, PT7, RBS, mcherry, fd terminator, sso, repts |
pGM1190 | tsr, aac(3)IV, oriT, to terminator PtipA, RBS, fd terminator, sso, repts |
pGM1202 | tsr, aac(3)IV, oriT, to terminator PtipA, RBS, C-terminal His6-tag, fd terminator, sso, repts |
pGM1192 | tsr, aac(3)IV, oriT, PSF14, RBS, N-terminal strep tag IIsc*, mcherrysc*, fd terminator, sso, repts |
pGM-Gus | aac(3)IV, oriT, PermE_gusA, repts (knock out vector) |
pGus21 | aac(3)IV, oriT, PermE_gusA, oripMB1 (knock out vector, non-replicative in Streptomyces) |
* Codon optimized for expression in Streptomyces