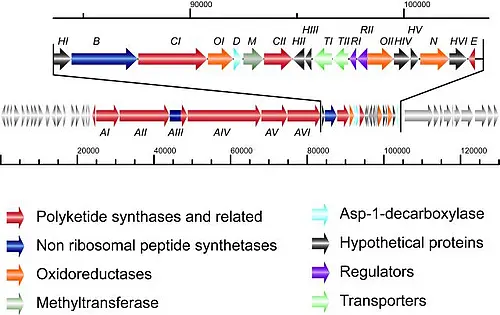
In our recent work, we identified and isolated the kirromycin biosynthetic gene cluster from a cosmid library of the producing strain Streptomyces collinus Tü 365 (Weber et al., 2003, J. Biotechnol. 106:221-232; Weber et al., 2008, Chem. Biol. 15:175-188). Based on genetic, biochemical and in silico data, 26 genes encoded on a 82 kb genomic fragment, were assigned to the kirromycin biosynthetic gene cluster (Figure 2).
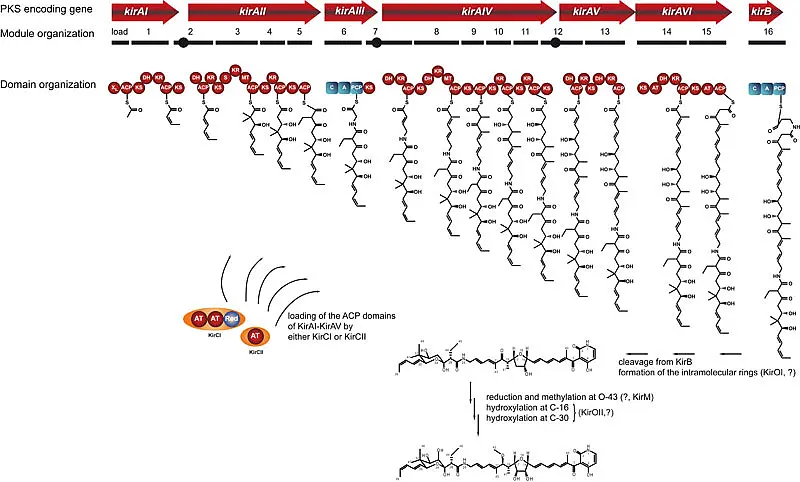
The carbon skeleton of kirromycin is synthesized by a unique combination of trans-AT polyketide synthases (PKS), cis-AT PKS and non-ribosomal peptide synthetases, which is subsequently tailored to yield bioactive kirromycin (Figure 3).
During our work, we were able to firstly identify a novel route to pyridone formation which involves the condensation of the non-proteinogenic amino acid β-alanine to an polyketide precursor. We showed, that β-alanine is not only provided by primary metabolism but also through an specific enzyme of the kirromycin biosynthetic gene cluster (Laiple et al., 2009, J. Antibiotics 62:465-468)
In contrast to many other PKS or NRPS gene clusters, the enzyme which is required to activate the carrier protein domains of the PKS/NRPS is encoded within the kirromycin biosynthetic gene cluster. We could demonstrate that the phosphopantetheinyltransferase KirP is able to convert both, the ACP and PCP domains of KirAI-AVI and KirB from their inactive apo-form into the active holo form (Pavlidou et al., 2011, FEMS Microbiol. Lett. 319, 26-33).