Shaker-like potassium channels in plants
Potassium is an essential macronutrient for plants (White et al., 2010); it constitutes the major inorganic, osmotic solute that drives cell expansion. It also regulates physiological responses towards environmental stresses. One of the most important functions and most studied is the opening and closing of the stomata which is achieved through turgor changes that follow K+-uptake and release. K+-transport through plasma membranes can either be facilitated by channels or secondary transporters.
Plants have evolved 3 different types of K+-channels, Kv-like, Tandem-Pore K+-channels (TPK) and the K+-inward rectifier (Kir) channels (Lebaudy et al., 2007). All three have orthologues in animals. The first channels identified through yeast complementation were members of the Kv-like family. The Arabidopsis genome encodes 9 Kv-like channels, of which 4 are inward rectifying (KAT1, KAT2, AKT1, SPIK), 2 outward-rectifying (GORK, SKOR), 1 silent (KC1) and 2 which are weak or unknown rectifying channels (AKT2/3, AKT5) (Pilot et al., 2003).
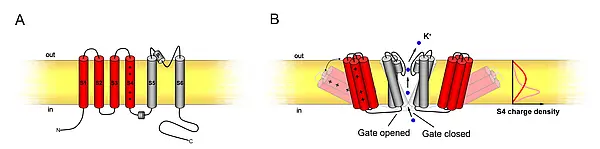
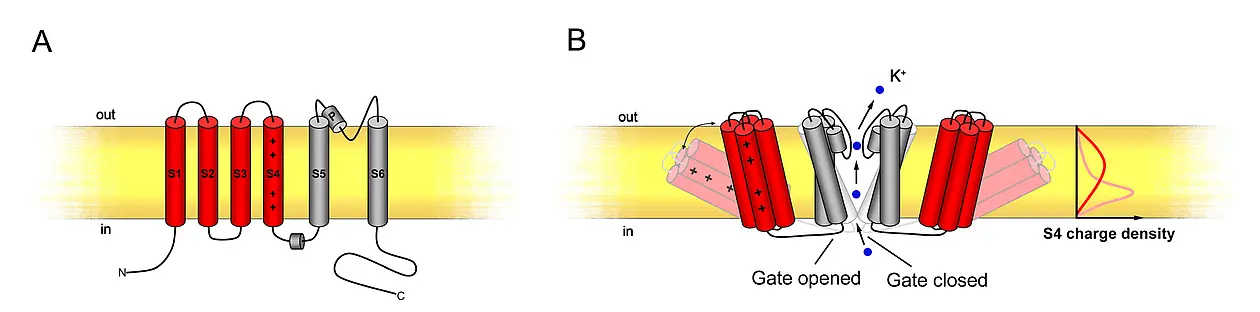
Each channel consists of 6 transmembrane α-helices (S1-S6) and incorporates a pore (P) loop between the S5 and S6 helices (see figure 6A). A functional channel is assembled by four α-subunits. These tetramers can consist of homomeric as well as heteromeric complexes adding another layer of regulatory potential to K+-transport (Dreyer et al., 1997; Lebaudy et al., 2007). The channels are voltage-gated; thus, they are regulated through changes in plasma membrane polarisation. The first four α-helices form the voltage sensor, the S4-domain containing regularly spaced, positively charged amino acids (arginine and lysine). The gate is formed as diaphragm between the base of the four S5-S6 helices, and voltage leads to a positional change of the voltage sensors which then draw open this diaphragm (see figure 6B).
{kind=link}