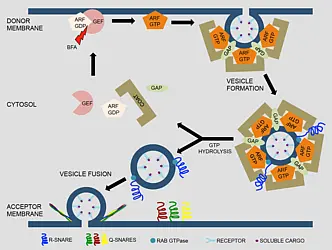
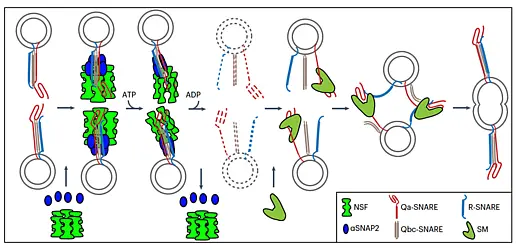
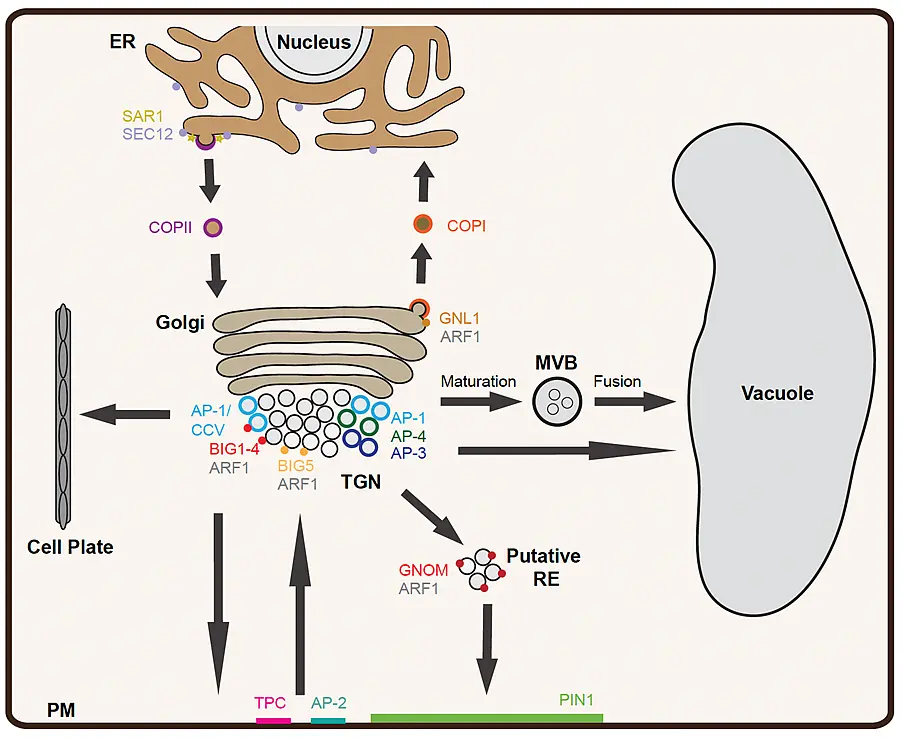
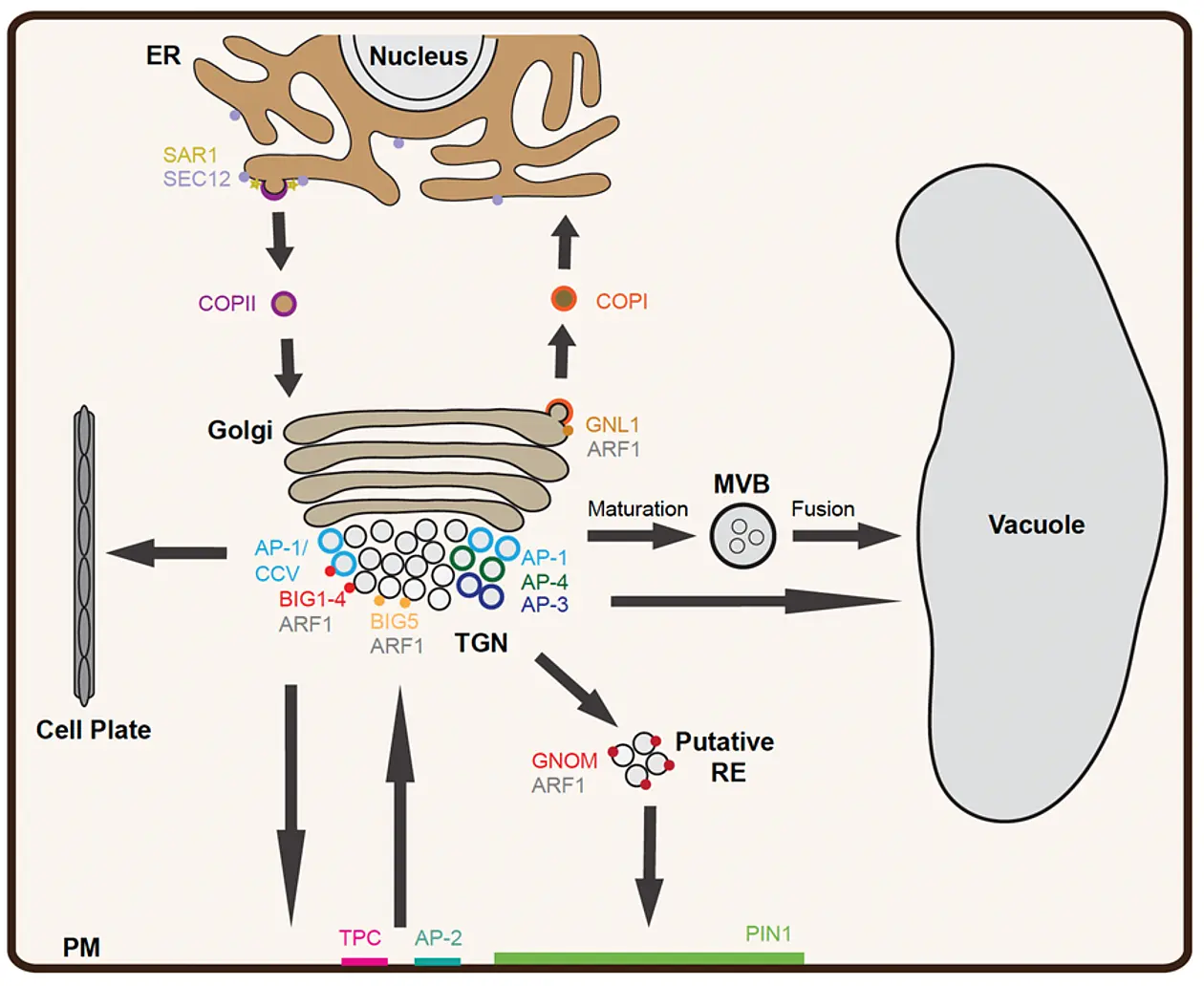
Figure 1: Membrane trafficking pathways in plant cells (schematic)
A secretory protein enters the endomembrane system at the ER and is transported to the Golgi using COP-II coated vesicles. At the ER exit site (ERES), SAR1 GTPase is activated by exchange factor SEC12 followed by recruitment of the COP-II coat complex. The retrieval of ER-resident proteins from the Golgi is achieved by retrograde Golgi-ER trafficking involving COP-I coated vesicles. The COP-I coat is recruited at the Golgi by ARF1 after its activation by ARF-GEFs, GNOM and GNL1. Transport of cargo proteins from cis-Golgi to trans-Golgi and from trans-Golgi to the TGN occurs via maturation. Transport from the TGN to the PM and to the cell plate is mediated by adaptin protein-1 (AP-1) complex/clathrin-coated vesicles (CCV) which are recruited by the activity of ARF-GEF BIG1-4 and ARF1. During endocytosis, vesicles are formed at the PM either by AP-2 or by TPLATE complex (TPC). The ARF-GEF BIG5 together with ARF1 regulates the endocytosis. The endocytosed proteins such as auxin-efflux transporter PIN1 are recycled back to the PM via the GNOM-positive putative recycling endosome (RE) in an ARF1-dependent manner or transported to the vacuole for degradation. The major vacuolar transport from the TGN involves maturation of the MVB from a sub-domain of the TGN followed by its fusion with the vacuole. In addition, other pathways mediated by AP-1, AP-3 and AP-4 also contribute to the post-Golgi-vacuole transport (modified from Singh and Jürgens, 2018)
{kind=link}