Research
Cell wall recycling metabolism in B. subtilis and E. coli
The bacterial cell wall consists of a huge, single macromolecule composed of polysaccharide strands crosslinked by oligopeptide, the so-called murein sacculus or peptidoglycan [Weidel,Pelzer 1964; Höltje 1998]. While the murein sacculus of Gram-negative bacteria is predominantly monolayered and accounts for less than 5% of the cell mass, the cell wall of Gram-positive bacteria is multilayered und usually makes up more than 20% of the cell mass [Seltmann,Holst 2002]. In both cases the sacculus has to be cleaved during cell growth (autolysis), to allow new material to be attached to or inserted in the existing stress-bearing cell wall (cell wall synthesis).
It was shown experimentally that E. coli and B. subtilis, representing the Gram-negativ and Gram-positive bacteria, respectively, break down their own cell wall during growth - a process that was termed "turnover" [Doyle,Koch 1987; Doyle et.al. 1988; Goodell,Schwarz 1985]. Massive turnover of about 50% of the cell wall per generation was long overseen by the fact that cell wall turnover products (at least in E. coli) are efficiently recycled. The complex peptidoglycan recycling pathway of E. coli is now about to be understood and a huge number of enzymes that are dedicated to the process of salvaging all components of the cell wall murein have been identified (Fig. 1).
The muropeptide recycling was first recognized by Goodell and Schwarz [Goodell,Schwarz 1985; Goodell 1985] and extensively studied furtheron primarily by James T. Park and coworkers [Park 1993; Jacobs et al. 1994; Park 1995]. However, it took until very recently that the recycling pathways for the amino sugars of the cell wall were elucidated [Uehara,Park 2004; Uehara et al. 2005; Uehara et.al. 2006; Jaeger,Mayer 2005]. My group has participated in the elucidation of the cell wall recycling pathways in the Gram-negative model organism E. coli (for recent reviews see Mayer, 2005; Mayer,Boos 2005; Jaeger,Mayer 2008). We elucidated the catabolic pathway for the cell wall sugar N-acetylmuramic acid (MurNAc), which involves transport and concomitant phosphorylation by the MurNAc-specific phosphotransferase system (PTS) transporter MurP (Dahl et al. 2004). The product of the MurP reaction, MurNAc 6-phosphate, is cleaved in the cytoplasm by a lactyl etherase (MurQ) representing a novel class of sugar-acting lyases/hydrolase (Jaeger et al. 2005; Uehara et al. 2006; Jaeger,Mayer 2008), which proceed through an unsaturated sugar intermediate (see below). MurQ is essential for the utilization of MurNAc from the environment and also for the recycling of the peptidoglycan of the own cell wall. The latter pathway involves the action of so-called lytic transglycosylases, which generate the 1,6 anhydro form of MurNAc, which is an intramolecular glycoside. MurQ is not only essential for growth on MurNAc but it is also required for utilization of 1,6-anhydromuramic acid (AnhydroMurNAc) derived either from the own cell wall (recycling) or imported from the medium [Uehara, 2005; Uehara, 2006].
References
Höltje, J.-V. (1998) MMBR, 62, 181-203.
Weidel, W., Pelzer, H. (1964) Adv. Enzymol., 26, 193-232.
Seltmann, G., Holst, O. (2002) The Bacterial cell wall, Springer, Berlin.
Doyle, R.J., Koch, A.L. (1987) Crit. Rev. Microbiol., 15, 169-222.
Doyle, R.J., et. al. (1988) Microbiol. Rev.. 52, 554-567.
Godell, E.W., Schwarz., U. (1985) J. Bacteriol. 162, 391-397.
Goodell, E.W. (1985) J. Bacteriol. 163, 305-310.
Park, J.T. (1993) J. Bacteriol. 175, 7-11.
Jacobs, C., et al. (1994) EMBO J. 13, 4684-4694.
Park, J.T. (1995) Mol. Microbiol. 17, 421-426.
Uehara, T., Park, J.T. (2004) J. Bacteriol. 186, 7273-7279.
Uehara, T., et al. (2005) J. Bacteriol. 187, 3643-3649.
Uehara, T., et al. (2006) J. Bacteriol. 188, 1660-1662.
Jaeger, T., et al. (2005) J. Biol. Chem., 280, 30100-30106.
Mayer, C. (2005) BioSepektrum 5, 626-619.
Mayer, C., Boos., W. (2005) EcoSal Chapter 3.4. Hexose/hexitol and pentose/pentitol metabolism, ASM press, Washington Online: www.ecosal.org
Dahl, U. et al. (2004) J. Bacteriol. 186, 2385-2392.
Jaeger, T., Mayer, C., (2008) Cell. Mol. Life Sci. 65., 928-939.
Adaptation of peptidoglycan catabolism in pathogenic and environmental bacteria
A renewed interest in the dynamics of the bacterial cell wall has emerged from recent discoveries in cutting-edge fields of molecular microbiology, for instance:
- bacterial cell cycle control, cell division, and morphology,
- bacterial heterogeneity, biofilm formation, and multicellular behaviour, and
- starvation response, dormant state entry, and resuscitation.
We address the role of the cell wall component murein (peptidoglycan) within this context by applying a systems biology approach to gain access to the “cell wallosome” at a whole. Our effort combines modern analytical, biochemical and genetic techniques. In particular, we will identify and quantify cell wall modifications and metabolites within cells adapted to certain growth conditions by LC-MS and genetic techniques, and we will identify novel enzymes and transporters using whole genome microarray analyses and 2D-electrophoresis/MS-fingerprinting and characterize them functionally and mechanistically using natural and synthetic substrates. Furthermore, we address gene regulation by determining transcription levels of target genes using quantitative PCR and reporter gene assays. In this way we will explore host adaptation of environmental and pathogenic microorganisms to improve our knowledge on the influence of cell wall metabolism on survival.
Structure and mechanism of MurNAc lactyl etherases (Carbon-Oxygen lyases)
Enzymes that cleave ether bonds are remarkable catalysts, because ethers (C-O-C), in general, are highly resistant to biodegradation due to their high bond energy and chemical inertness [White, 1996]. As a consequence, ether compounds frequently accumulate in the environment including most problematic pollutants. In fact, this relative inertness may be the reason that nature sometimes makes use of the ether bond, e.g. in recalcitrant substances like the lignin of wooden plants [Masai, 2007]. The trivial name etherases classifies enzymes that catalyze the scission of ether bonds by exhibiting a wide variety of mechanisms:
- oxygenative cleavage via monooxygenases;
- oxidation of the carbon atom α-linked to the ether bond, followed by hydrolysis of the resulting ester;hydroxyl shift mechanisms;
- direct hydrolysis of the C-O bond,
- anaerobic cleavage of methyl-aryl ethers,
- oxidative mechanisms,
- reductive mechanisms
and finally - carbon-oxygen lyase-mediated cleavage [White 1996].
MurNAc etherases catalyze a lyase-type reaction [Jaeger 2005]; β-elimination of the lactic acid alkoxyl group at C3 is follwed by hydration of the unsaturated sugar intermediate. Usually, an alkoxyl group is not a leaving group, and so the lacyl ether alkoxyl substituent of MurNAc might be protonated prior to cleavage (Fig. 2). Unlike other ether compounts the lactyl ether substituent of MurNAc is rather unstable under alkaline conditions. At pH > 12 the proton (H2) α-positioned to the carbonyl of the sugar is readily eliminated. Likewise the enzyme might provide a basic residue that catalyzes the same deprotonation reaction. Hence, a least two catalytic residues are required: a general base that removes the H2 and an acid catalyst that protonates the leaving group. The acidity of the H2 is greatly increased in the open chain form of the sugar, hence a ring opening step, like in other other sugar phosphate isomerase (SIS) family proteins is suggested. The unsaturated phosphosugar intermediate, which is released by the elimination of lactic acid has a considerable lifetime and can be detected either by a colorimetic assay (Morgan-Elson assay) or by mass spectrometry. In the Morgan-Elson reaction the very same unsaturated amino sugars are generated by heating (boiling for 3 min) under alcaline conditions (borate buffer pH > 10). This step can be substituted by an incubation with etherase. The second part of the Morgan-Elson reaction is the reaction of Ehrlich reagent (dimethylaminobenzaldehyde, DMBA in acidic acid) which leads – in both cases - to to the formation of a violet colored adduct.
The intermediate of the etherasse reaction can be visualized in particular, when the reaction is run „in reverse“, indicating that an equilibrium of intermediate and product is formed; in the forward reaction the intermediate is readily converted into product by hydration of the double bond. Possibly, the same catalytic amino acid residues are required for the hydration step as for the elimination step: the former acid catalyst would then functions as a base deprotonating the incoming water molecule and the protonated former general base provides its proton. In priciple, MurNAc etherases should be able to operate in reverse, provided that a good acceptor is present. With this enzymes the synthesis of a range of amino sugars with C3-modifications should be a possible.
References
Jaeger, T., et al. (2005) J. Biol. Chem., 280, 30100-30106.
Masai, E., Katayama, Y. and Fukuda, M. (2007) Biosci. Biotechnol. Biochem. 71, 1-15.
Mayer, C. (2005) BIOspektrum 5, 616-619.
White, G.F., Russell, N.J. and Tidswell, E.C. (1996) Microbiol. Rev. 60, 216-232.
Structure and mechanism of regulators of MurNAc metabolism
E. coli utilizes the bacterial cell wall sugar N-acetylmuramic acid (MurNAc) dependent on the etherase MurQ, which catalyzes the cleavage of the lactyl ether substituent of MurNAc-6-phosphate. High-level expression of murQ is essential for growth on MurNAc as the sole source of carbon and energy, whereas low-level constitutive expression is sufficient for the recycling of peptidoglycan fragments continuously released in small quantities from the cell wall during growth. Here we characterize for the first time the expression of the murQ operon and its regulation by MurR, a member of the poorly characterized RpiR/AlsR family of transcriptional regulators. murR and murQ are expressed from non-overlapping face-to-face (convergent) promoters, yielding transcripts that are complementary in their 5'ends. MurR forms a stable multimer (most likely a tetramer) and presumably binds as such to two adjacent inverted repeats within an operator region. In this way MurR represses transcription from the murQ promoter and also interferes with its own transcription. MurNAc-6-phosphate, the substrate of MurQ, is the inducer and weakens binding of MurR to the operator. Furthermore, high-level murQ transcription depends on activation by cyclic AMP (cAMP)-catabolite activator protein (CAP) bound to a class I site upstream of the murQ promoter (Jaeger,Mayer 2008).
Engineering carbohydrate-active enzymes and regulators
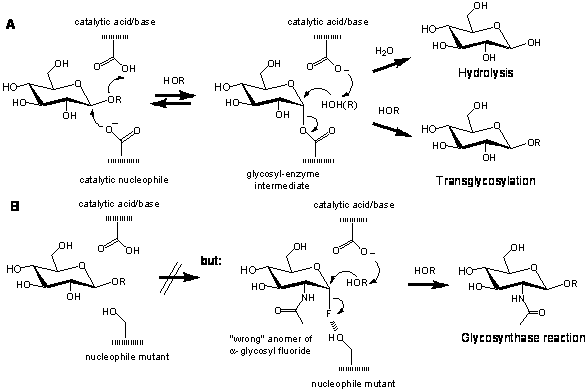
Glycosyl hydrolases modified at the catalytic nucleophile position (glycosynthases) can be used for the synthesis of oligosaccharides. They act differently from the wild-type enzymes in that they are incapable of hydrolyzing glycosides but perform transglycosylation reactions when presented with the “wrong” anomer of the glycosyl fluoride substrate along with a suitable acceptor (Fig. 3). Specific products, high yields and a simple one-step reaction from cheap substrates are the advantages of enzymatic synthesis with glycosynthases. Pasted Graphic.pict ¨ Fig. 3. Mechanism of Agrobacterium faecalis β-glucosidase modified at the catalytic nucleophile (AbgE358S) acting as a glycosynthase (A). The glycosynthase performs transglycosylation reactions when presented with the “wrong” anomer of the glycosyl fluoride substrate along with a suitable acceptor (B). The ‘original’ glycosynthase, derived from Agrobacterium sp. β-glucosidase (Abg) by mutating the nucleophile glutamate to alanine (E358A), synthesizes oligosaccharides in yields exceeding 90 % [Mackenzie 1998]. This mutant was re-cloned with a His6-tag into a pET vector, allowing gram scale production and single step chromatograpic purification [Mayer et al. 2000]. A dramatic, 24-fold, improvement in synthetic rates was achieved by substituting the nucleophile with serine, resulting in improved product yields, reduced reaction times and an enhanced synthetic repertoire [Mayer et al. 2000; Tolborg et al. 2002]. Thus poor acceptors for Abg E358A, such as pNP-GlcNAc, are successfully glycosylated by E358S, allowing the synthesis of pNP-β-LacNAc. The increased glycosylation activity of Abg E358S likely originates from a stabilizing interaction between the Ser hydroxyl group and the departing anomeric fluorine of the α-glycosyl fluoride (Fig. 3B). The nucleophile mutants can also catalyze nucleophilic halogenation [Zechel 2001]. The enzymic synthesis of carbon-fluorine bonds is rarely used in nature. We developed a fast and easy screening procedure for the discovery of efficient new glycosynthases. Using a two plasmid system, this coupled enzyme screen can be used on agar plates to select improved glycosynthases from mutant libraries. These libraries were generated by a PCR-based site/region-directed mutagenesis In a screening experiment using a library of saturated mutants at the catalytic nucleophile position/region. Abg E358G was identified using this screen, which is the so far best Abg glycosynthase that has a turnover rate for the carbon-fluorine bond cleavage that exceeds 2 s-1 [Mayer, 2001; Zechel, 2001; Kim, 2004].We are now trying to adapt the glycosynthase strategy for the MurNAc etherase ([Jaeger, 2005]; see above LINK). If this works it would be possible to generate N-acetylglucosaminide derivates specifically substituted at position C3.
References
Jaeger, T. et al. (2005) J. Biol. Chem., 280, 30100-30106.
Kim et al. (2004) J. Biol. Chem. 279, 42787-93.
Mackenzie, L., et al. (1998) J. Am. Chem. Soc. 120, 5583 - 5584.
Mayer, C. et al. (2000) FEBS Lett. 466, 40-44.
Mayer, C. et. al. (2001) Chem Biol. 8, 437-443.
Tolborg et al. (2002) J. Org. Chem. 67, 4243-4249.
Zechel et al. (2001) J. Am. Chem. Soc. 123, 4350-4351.