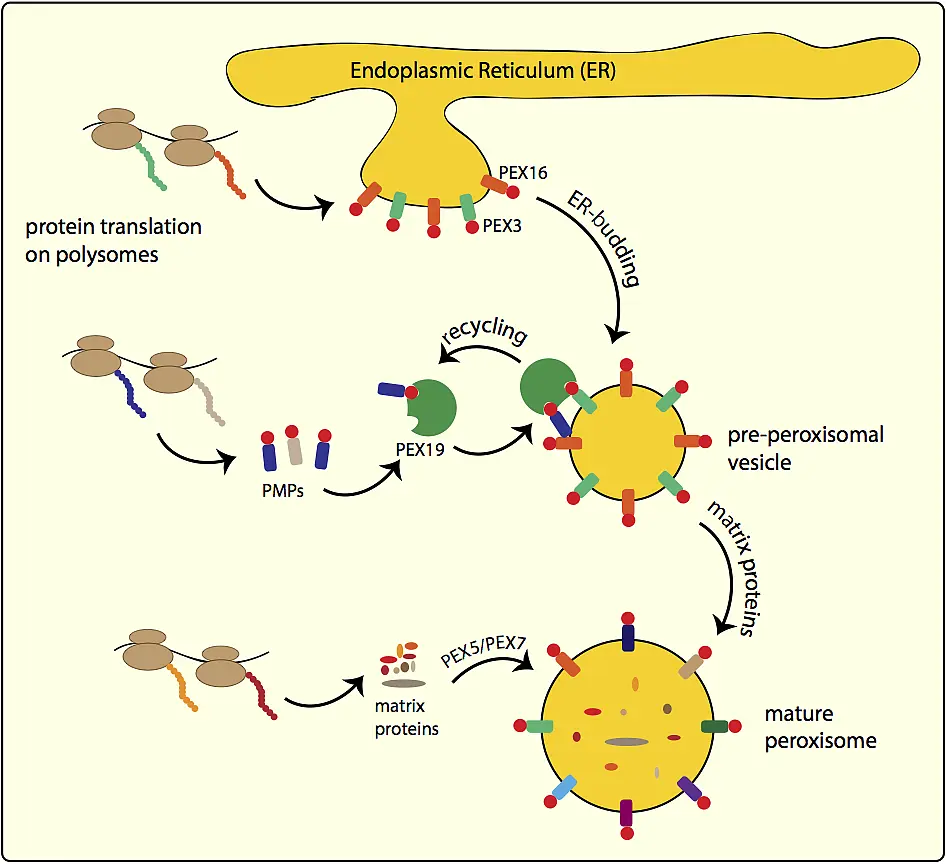
The early peroxins PEX3, PEX16 and PEX19 are also essential for posttranslational insertion of peroxisomal membrane proteins (PMPs) into the peroxisomal membrane. Most PMPs have a quite variable membrane targeting sequence (mPTS) and are bound in the cytosol after translation by their receptor PEX19. The cargo-loaded PEX19 binds to the import-receptor PEX3 at the peroxisomal membrane and the PMP is imported. Afterwards PEX19 is recycled into the cytosol for another PMP import cycle.
We were able to crystallize the cytosolic part of PEX3 in combination with N-terminal parts of PEX19 and could therefore improve the understanding of the PEX3-PEX19-PMP binding mechanism. Structural analysis revealed three highly conserved patches within PEX3: The PEX19-binding region on top of a α-helical bundle, a hydrophobic groove potentially involved in PMP binding and an obviously non-functional acidic cluster.