Cognitive Neurology
Overview

{kind=link}
The group addresses a variety of topics that, at first glance, may look disparate. Yet, there is a common denominator, namely the manifold consequences of the fact that we dispose of motricity — motricity, arguably the ultimate reason for the development of brains. We are interested in the question of how our brain is able to plan and to execute optimal goal-directed movements and how sensory systems are able to deal with the influence of movement on the sensory signals, ensuring movement independent percepts. And we deal with the information we take from the movements made by others in order to understand their needs and intentions, allowing us to select appropriate behavioral responses. And whatever perspective we take on movement, we always try to understand what it means to suffer from disturbances of the systems involved.
Research topics
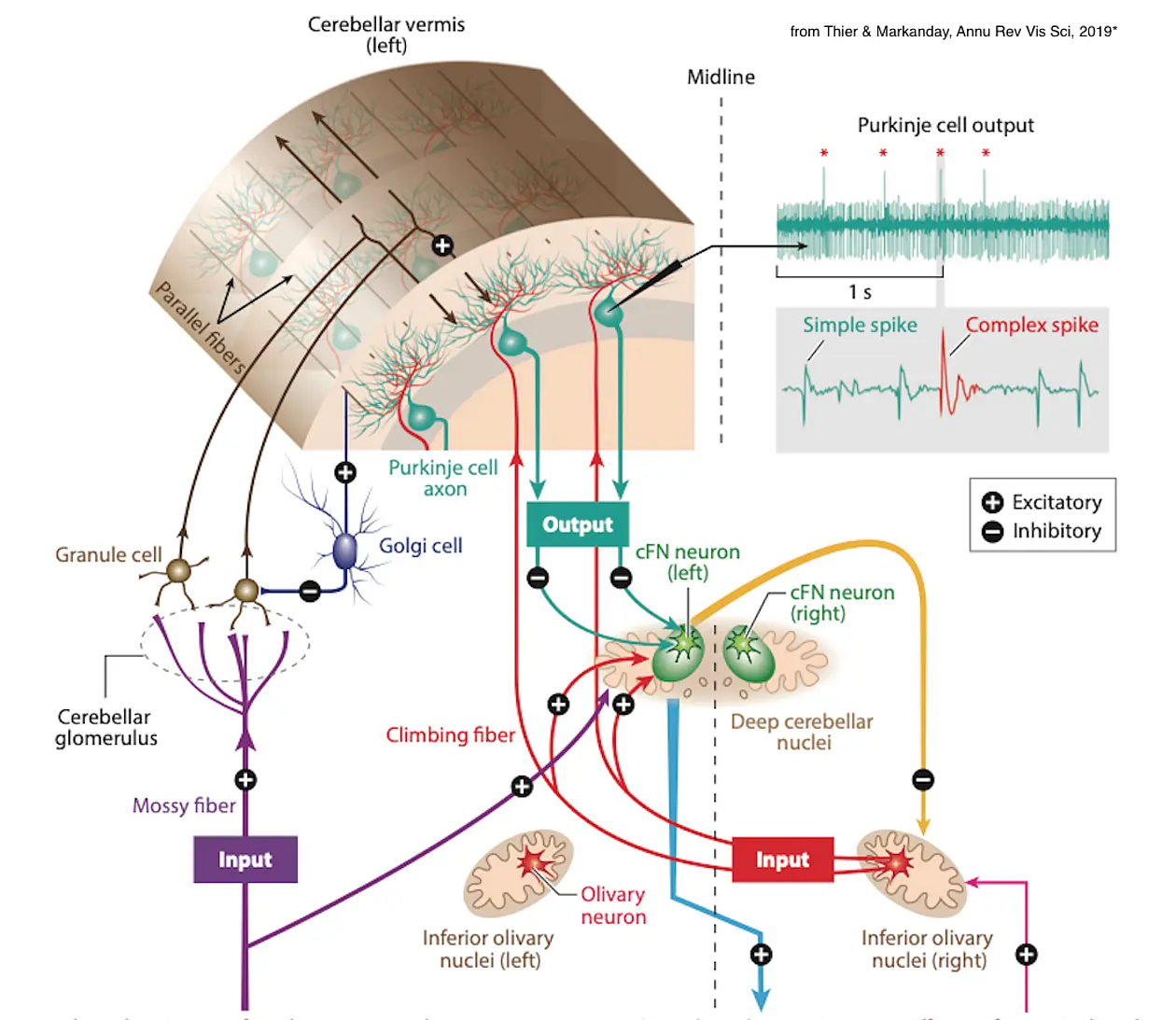
Cerebellar Control of Movement
The core symptom of cerebellar disease is ataxia, the inability to carry out movements precisely and reliably. The pathophysiology of ataxia, or dysmetria, has as yet escaped proper understanding. This is a direct consequence of the many unanswered questions on the normal physiology and function of the cerebellum Neurology has seen numerous attempts to mitigate cerebellar ataxia and related problems. Whereas the usually poorly justified pharmaceutical approaches were bound to fail, rehabilitation training has time and again shown to be beneficial, in particular, when insights on the specifics of cerebellum based motor learning were taken into account. However, the potential of rehabilitation training is limited. If we strive for more substantial therapeutic options, we will have to better understand the normal working of this neuronal machine. This need will have to be met, independent of whether we dream of replacing the lost machine by technical means, i.e. a cerebellar prosthesis, or by regenerating circuits using cell biological approaches. Whichever of these two science fiction approaches may one day materialize, it will have to reflect the blueprint of the normal neuronal machine. We have worked on this blueprint for many years, in the early years focusing on the pontine nuclei, the major interface between cerebral cortex and the cerebellum.
For review see:
*Thier P, Markanday A (2019) Role of the vermal cerebellum in visually guided eye movements and visual motion perception. Annu Rev Vis Sci 5:247-268; doi: 10.1146/annurev-vision-091718-015000. Epub 2019 Jul 12.
Dash S, Thier P (2014) Cerebellum-dependent motor learning: lessons from adaptation of eye movements in primates. Progr Brain Res 210: 121-155.
Trying to unravel the underpinnings of social interactions
In order to interact successfully with others we have to develop a theory of mind (TOM), a concept of the other’s interests, desires and goals. Exploiting non-verbal information offered by the other’s body is a key step in developing a TOM: The gaze of the other allows us to identify her/his objects of attention, to establish joint attention and to project our own object-associated needs and aspirations onto the other one. Additional information on the needs of the other is provided by the actions directed at these objects of joint attention.
Both disturbances of gaze following and of action understanding have been suggested to underlie human autism. The term ASD (autism spectrum disorder) or simply autism tries to embrace a continuum of developmental disturbances, which lead to impaired social interaction and communication skills, the inability to establish normal relationships, reduced language skills, repetitive and stereotypic behaviors, in many cases to intellectual disability and a large variability of possible complicating secondary symptoms. Asperger syndrome and high functioning autism, terms which are often used synonymously, can be understood as mild variants of autism in which disturbances of social skills may come with no or little deficits in other domains and gradual transition to a normal personality with distinct traits. Subjects with autism lack spontaneous eye-gaze following under natural conditions. As gaze following and joint attention based on it are major building blocks of a viable TOM, this gaze following alteration could easily explain the unavailability of a normal TOM in autism. However, subjects with autism, at least the older ones, are able to shift attention based on eye-gaze cues and they may exhibit even faster orienting guided by eye gaze. In collaboration with the Tübingen Department of Psychiatry we have launched psychophysical and BOLD-imaging studies of adult patients suffering from high functioning autism in order to test specific hypotheses suggested by our recent psychophysical work on monkeys and healthy human subjects.
Exemplary publication
Ramezanpour H, Thier P (2020) Decoding of the other´s focus of attention by a temporal cortex module. Proc Natl Acad Sci USA, doi: 10.1073/pnas.1911269117. [Epub ahead of print].
Motricity-invariant visual perception
We dispose of motricity, allowing us to explore the world and to influence the world. The downside of motricity is its pervasive influence on sensation. It forces our brains to deal with the question if sensory signals are a consequence of motricity or, alternatively, a result of an event in the world. Only the latter is ecologically relevant and therefore should lead to a veridical percept. Our work, has provided compelling evidence that our ability to perceive visual motion as it unfolds in the external world is a consequence of the fact that we are able to generate a reliable prediction of the expected visual motion consequences of our eye movements, subtracted from the visual motion input as seen by the eyes. We could show that this prediction is contributed by the cerebellum, continuously fine tuning a first, crude sketch of the necessary prediction provided by supplementary motor cortex. The early stages of the cortical processing of visual motion are ignorant of the source of visual motion being external or due to the eye movement. The optimized prediction coming from the cerebellum enters the cortical motion hierarchy only at the level of the newly discovered area VPS in the caudal lateral fissure, where motion in the external world is extracted, taking the predicted motion consequences of the eye movements, coming from the cerebellum into account. Lesions of this region and in general, a dysfunction of this inferential mechanism is - as we could show — a major cause of dizziness.
In our most recent work, we have turned to the related problem of orientation stability, our ability to perceive the visual world to be upright, although tilting the head relative to the gravitational vector may cause substantial tilting of the retinal image.
Exemplary publication
Daddaoua N, Dicke PW, Thier P (2014) Eye position information is used to compensate the consequences of ocular torsion on V1 receptive fields. Nature Communications. 5:3047. doi: 10.1038/ncomms4047