Transcranial Magnetic Stimulation (TMS) is a technique that enables stimulation of the human brain non-invasively with limited discomfort for the subject. It is based on the principles of electromagnetic induction. By applying a brief pulse of current through a stimulation coil held on the subject’s scalp, a time-varying magnetic field perpendicular to the current is produced. In turn, this magnetic field induces an electrical current in the brain. Depending on the stimulation parameters, the current induced will transiently interfere with cortical activity, which can result in altered behavior.
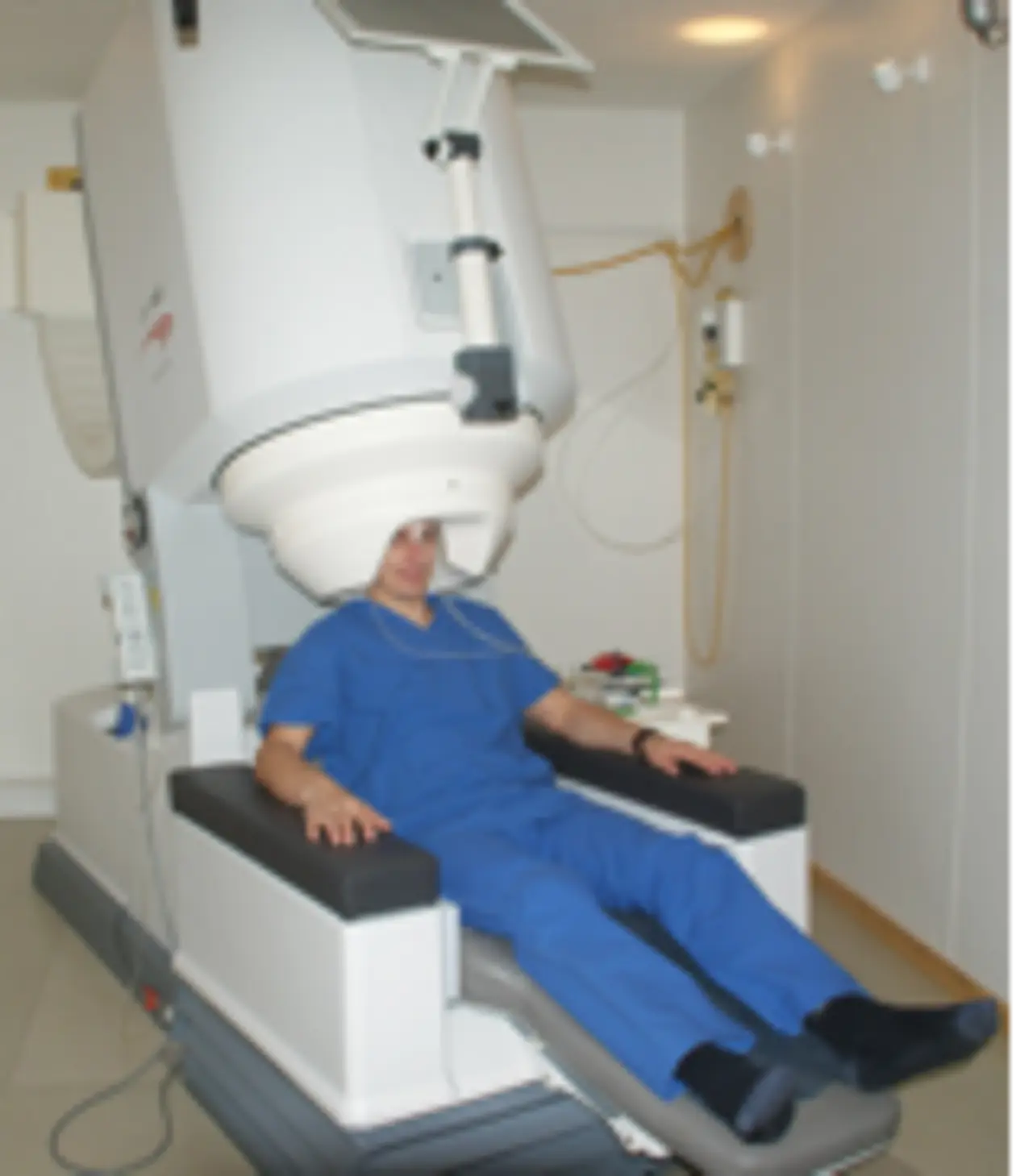
The TMS equipment consists of a pulse generation unit, the stimulator, and an electromagnetic stimulation coil. The most widely used types of coil are the circular coil and the figure-of-eight coil. Figure-of-eight coils can be approximately viewed as two round coils mounted side to side, with the current rotating in opposite directions in the two coils. While with circular coils the maximum electric field induced in the brain lies in an annulus under the coil, the electric field with figure-of-eight coils is strongest under the centre of the coil. Figure-of-eight coils thus maximize the focality of TMS.
Another important feature of TMS is that the induced magnetic field is inversely proportional to the square of the distance between the coil and the cortex, meaning that only superficial neural structures are sensitive to direct stimulation, whereas deeper areas might only be excited indirectly by propagated activity from the region beneath the coil. TMS can be applied as single pulses or as train of pulses (rTMS). The effects of single TMS pulses are short-lived and several seconds are needed before the next pulse can be delivered. Conversely, rTMS can modulate the excitability of the stimulated area even beyond the duration of the TMS application with the specific effect depending on the prior history of activation.
Despite the extensive use of TMS, the exact mechanisms still need to be defined. Two main mechanisms have been proposed. The virtual lesion approach assumes that the application of a TMS pulse temporarily lesions or disturbs the function of a population of neurons. Alternatively, TMS may induce neural activity by adding ‘extra activity’ to ongoing processes. Both mechanisms will alter neural processing and may manifest themselves in behavioral changes such as longer/shorter responses, errors, lowered detection threshold. Yet, TMS may not only change local neuronal activity directly, but also affect processing in remote brain regions either via neural interregional connections or due to compensatory mechanisms.
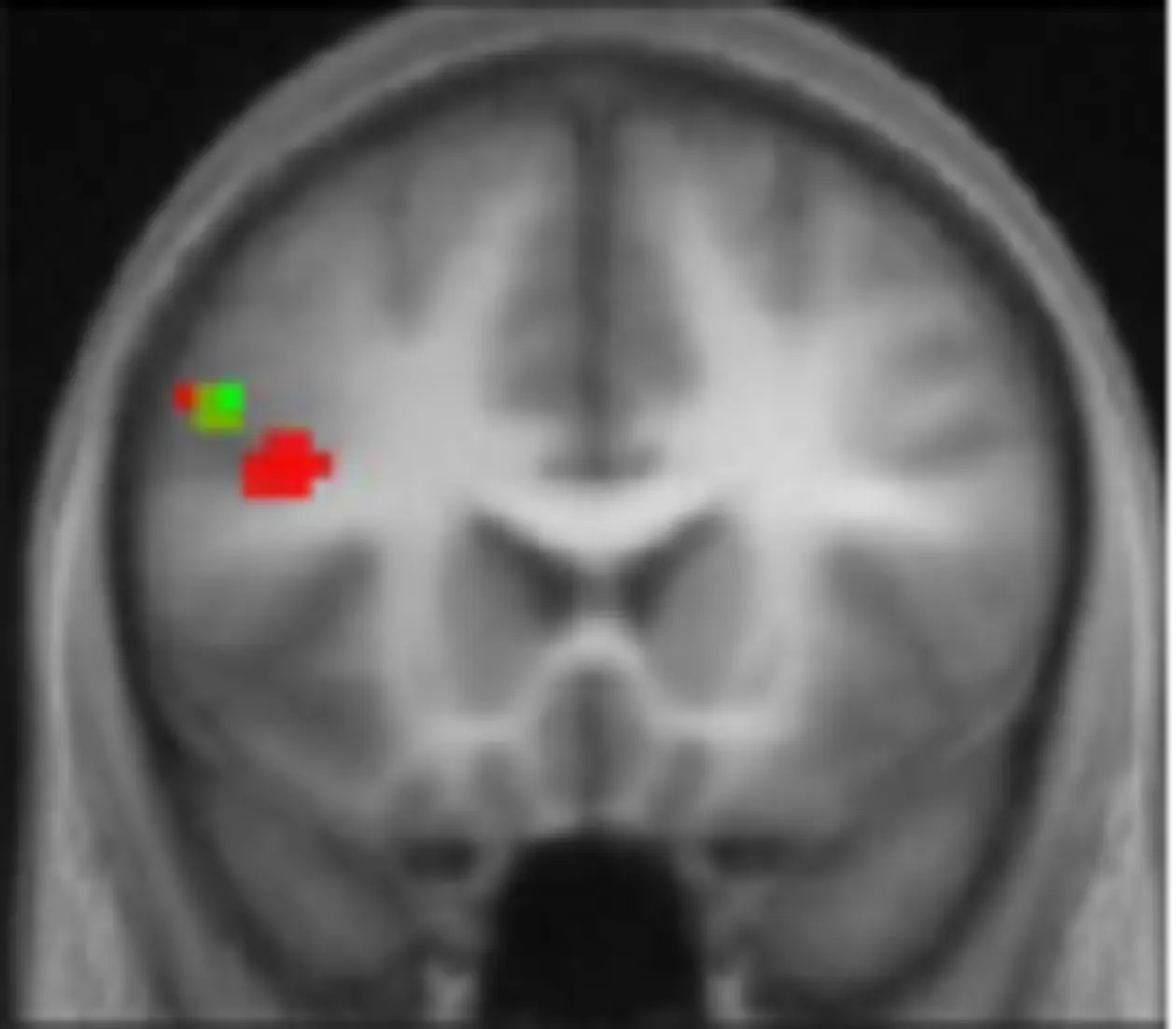
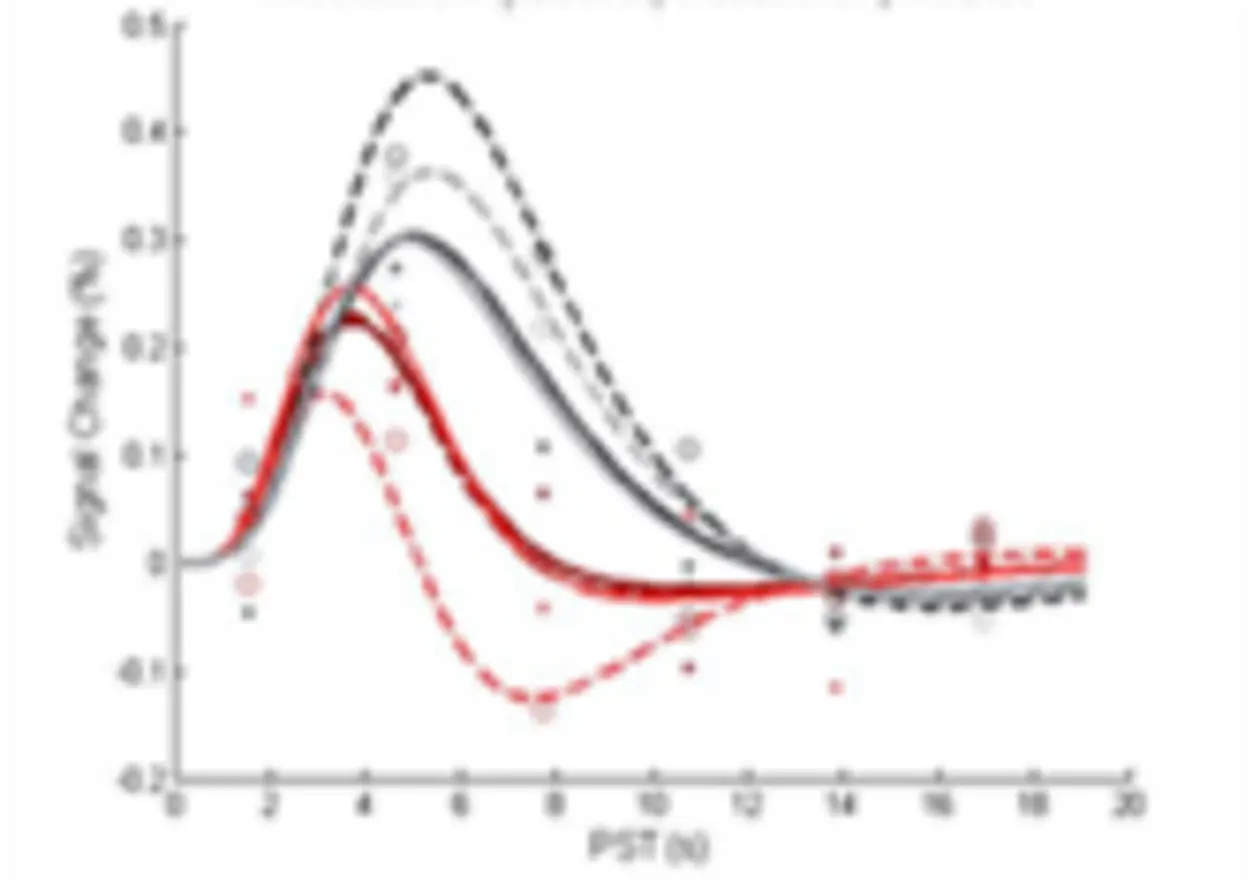
Such remote effects are not revealed by standard TMS studies, where inference is usually limited to the targeted site of stimulation. Instead, the combination of TMS and fMRI (e.g. interleaved TMS & fMRI) allows one to monitor the effects of TMS on neural activity directly not only at the site of stimulation but also within the entire brain, allowing also for inferences about the connectivity between the stimulated and co-activated areas.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}