17.09.2020
A Butterfly’s Wingbeat: The Tübingen Root of PGRMC1 Research
In Conversation with Research Alumnus Dr. Michael Cahill, Lecturer in Biochemistry and Cell Biology at Charles Sturt University in Wagga Wagga, Australia
What is your connection with Tübingen and the University?
I obtained a PhD in Pathology specializing in molecular cell biology, transcriptional regulation and signal transduction in Alfred Nordheim’s lab in Hannover. From 1997-2000 as a young postdoc I started a proteomics lab in Alfred Nordheim’s new Institute for Molecular Biology at the University of Tübingen [1]. Today, that activity has given rise to the Proteome Centre Tübingen, which is still in the same building. But I had moved on before its foundation. Together Alfred and I were scientific co-founders of ProteoSys AG, where from 2000-2007 I managed technology development and all cancer research.
My recent research has pursued characterization of signaling surrounding the protein Progesterone Receptor Membrane Component 1 (PGRMC1), which we discovered to be differentially phosphorylated while working at ProteoSys, in a productive collaboration with the Tübingen Frauenklinik [2-4], who later extended the collaboration with the group of Xiangyan Ruan in Beijing [5-7]. The startup funding for ProteoSys was provided by Tübinger Seed Fonds KG (Seed) in 2000, so my current research results are directly dependent upon their activity, and of course upon Alfred Nordheim’s faith in letting me dabble in proteomics in his new lab, which was untried territory for him. Thank you Seed and thank you Alfred Nordheim.
Where are you based?
Altogether I worked in Germany for 18 years including my time in Tübingen. I now work in the School of Biomedical Sciences at Charles Sturt University in Wagga Wagga, NSW, Australia. My current expertise is still in the interpretation of omics results and biological syntheses, as exemplified by recent [8, 9] and previous [2, 3] publications.
PGRMC1 phosphorylation
The system that regulates PGRMC1 phosphorylation is a major driver of cell function through altered signaling activity and epigenetics. My group has recently demonstrated that PGRMC1 phosphorylation status can induce dramatic changes in not only cell metabolism, morphology, and cytoskeleton/mitochondria, but also CpG genomic methylation [8, 9]. I believe that this is important for things like early embryology, neurodifferentiation and synaptic function, differentiation status, and germline/soma segregation, all of which permeate normal and pathological adult cell functions. I have newly shown that PGRMC1 affects mitochondrial form and function and also modulates PI3K/AKT signaling [8], which may relate it not only to cancer and AD etiology, but also potentially to ageing processes through insulin/insulin-like growth factor-1 (IGF-1) signaling.
PGRMC1 is a cytochrome b5 fold protein
PGRMC1 is a cytochrome b5 fold protein and the archetypal member of the membrane associated progesterone receptor (MAPR) family, which is characterized by an insertion on the surface of the cytochrome b5 fold which I call the MAPR interhelical insertion region (MIHIR) [10-12]. PGRMC1 interacts with lanosterol demethylase (cytochrome P51A1) [2] to catalyze the 14-demethylation of lanosterol from yeast to mammals [13, 14]. One fun fact is that this is the most conserved cytochrome P450 reaction in eukaryotes, being present in all kingdoms [15] (and therefore probably stemming from the first eukaryote).
There is an alternative competing reaction in some mammalian tissues, but the direct product of the CYP51A1 reaction is follicular fluid meiosis-activating sterol (FF-MAS) [16]. Meiosis, and the related mitosis, had to be invented by the first eukaryotes. Because this is the very first steroidogenic reaction after lanosterol is cyclized from the mevalonate pathway, the steroidogenic reaction should have been one of the first to evolve in eukaryotes. The strong phylogenetic conservation of the catalyzing proteins PGRMC1/CyP51A indicates its importance for eukaryotes. The steroidogenic and MIHIR-associated functions of PGRMC1 seem to have arisen new in eukaryotes and were probably important in eukaryogenesis and the regulation of mitochondrial function.
PGRMC1 membrane trafficking seems both associated with the MIHIR and acquired by PGRMC1-like proteins early in eukaryotic evolution. I’ve recently shown that this region of PGRMC1 contains a protein interaction motif similar to many myosins [12]. We [17] and others [18] have shown that PGRMC1 interacts with components of the actin cytoskeleton [19]. This is probably associated with the motive force required for regulated exo- and endo-cytosis (reviewed by: [20]). I think that this function was originally required in eukaryogenesis to produce and transport steroids to the mitochondrial membrane in response to oxygen (unpublished), in a world where the endosymbiotic proto-mitochondrion detoxified environmental oxygen [21]. PGRMC1 still modulates mitochondria [8], and appears to direct root level eukaryotic programming (analogously to being able to circumvent older Microsoft Windows security systems via a DOS-level entry point).
The much later origins of the animal embryological gastrulation organizer (e.g. the Spemann-Mangold organizer of Xenopus, and Tübingen) and subsequent tissue differentiation mechanisms seem linked to the acquisition of PGRMC1 tyrosine phosphorylation. This appeared concurrently with gastrulation in the last eumetazoan common ancestor (LEUMCA) that gave rise to cnidarians and bilaterally symmetrical animals [12], and seems to have imposed sophisticated levels of regulation onto ancient processes.
Mutation
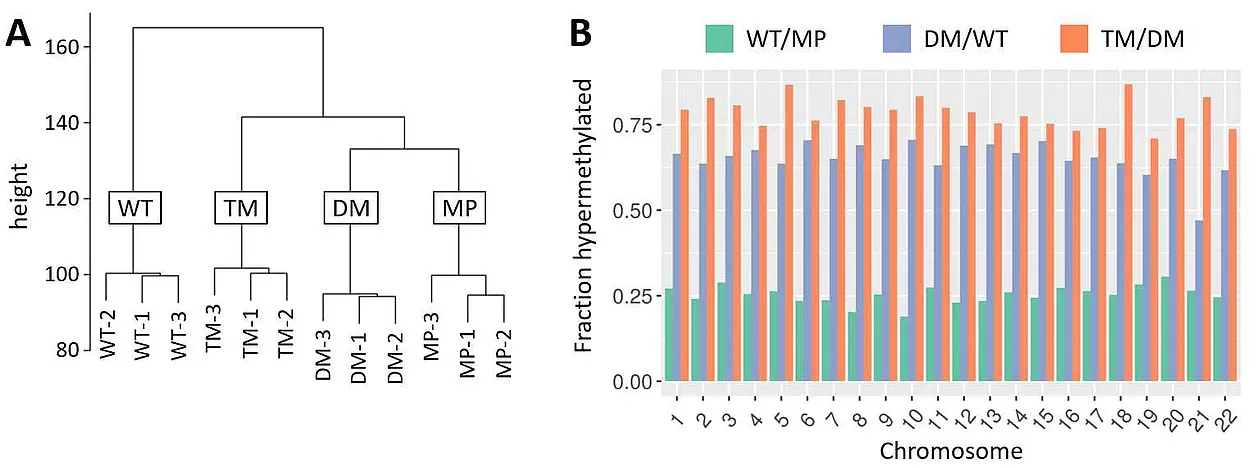
Mutation of one of the PGRMC1 phospho-tyrosines (Y180) in human cell culture caused dramatic cell morphological changes and energy metabolism protein abundance changes that resembled insulin/glucagon activity and altered glycolysis [8] (PGRMC1 is associated with diabetic biology [22, 23]). These changes were associated with dramatic epigenetic CpG genomic methylation changes (Figure 1), inducing a hypermethylated state reminiscent of embryonic stem cells, and potentially resembling the state of the pre-gastrulation LEUMCA genome. Each phosphorylation mutant state of the Y180 regulatory motif in cell culture induced a separate distinct fingerprint of genomic methylation [9], revealing PGRMC1 phosphorylation to be a previously unrecognized major driver of animal differentiation status. Y180 was also required for efficient tumorigenesis in mice [8]. An urgent need clearly exists to characterize this system.
PGRMC1 has long been known to operate in neurons. Cognition Therapeutics Inc. (CogRx, Pittsburgh) are developing a curative drug for AD which has given me valuable insights into the biology involved. Since 08/2013 I have served on their Scientific Advisory Board because PGRMC1 turns out to be at the center of the mechanism of their anti-Alzheimer’s drug. In June 2020 CogRx were awarded a $75.8 million Alzheimer’s Clinical Trials Consortium (ACTC) grant from the National Institutes of Health (NIH), the largest ACAT grant to date.
The novel roles for PGRMC1 which I recently discovered place it at a central point in animal evolution and the maintenance of cellular differentiation status. This newly emergent biology encompasses the amyloid beta and Tau perturbations associated with AD, which are linked by PGRMC1 membrane trafficking of the membrane receptors as the LDLR of ApoE4, in conjunction with the Sigma-2 receptor TMEM97 [24]. A protein complex involving PGRMC1 and the Sigma-2 receptor is targeted by the CogRx synaptorestorative drug CT1812 (in clinical Phase 2) that reverses phenotypes of AD such as Tau phosphorylation in human patients and cognitive impairment in rodent models. PGRMC1-mediated membrane trafficking was required for the toxic accumulation of amyloid beta oligomers on synapse surfaces as part of CT1812 drug mechanism [25]. It has long been known that PGRMC1 interacts with deleted in colorectal carcinoma (DCC) to mediate embryological axon guidance [see references in: 2]. Netrin signaling via DCC is essential in long term potentiation (LTP) [26], which is defective in AD. The simplest interpretation of the mechanism of action of CT1812 is that it prevents PGRMC1 from correctly interacting with DCC in synapses to enable synaptic plasticity.
Not only is PGRMC1 biology aligned with the above major ‘usual suspects’ of AD (Abeta, tau, ApoE4), but its newly emergent biology is also aligned with other known aspects of AD. We showed that PGRMC1 exerts dramatic influence over glucose metabolism, epigenetic status and mitochondria [8, 9], all of which are perturbed in AD neurons [27-29]. The PGRMC1 target complex of CT1812 is involved with ApoE/lipoprotein internalization of Aβ oligomers [30] that cause AD. Another AD marker, Tau pathology, is linked to ApoE [31, 32] so that the PGRMC1 complex is associated with most major markers of AD pathology. My hypothesis is that Abeta oligomers bind to the PGRMC1 complex, preventing normal PGRMC1 function and leading to AD. i.e. PGRMC1 biology causes AD! Interestingly, PGRMC1 is involved in the diurnal buildup of Abeta oligomers in synapses by day, and their clearance during sleep which is required for memory formation [33], strengthening the link between PGRMC1 and cognitive neural functions. My future research will hopefully examine the roles of PGRMC1 biology in CT1812’s mechanism of action on AD, and the role of PGRMC1 in cognition.

{kind=link}
Teilen




